
Кортексин® для детей | ГЕРОФАРМ
Этот термин часто встречается в медицинских публикациях, СМИ, в рекламе лекарственных препаратов. Возможности нейропротекции заложены в самой природе мозга, в генах, на уровне регуляторных нейропептидов. Суть нейропротекции состоит в том, что лечебный процесс способствует не только защите пострадавшей группы нейронов, но и обеспечивает ее дальнейшее функционирование. Для медицины важен вопрос — существуют ли адекватное фармакологическоое воздействие, способное запускать эти природные механизмы и поддерживать их на необходимом уровне? В этой связи, поиск, создание и апробация новых фармацевтических средств являются и будут являтся одним из наиболее важных направлений современной фармакологии.
Очевидно, что поиск новых нейропротекторов представляет собой сложный процесс, требующий объединенных усилий врачей, биологов, фармакологов на всех этапах. В этом отношении особого внимания заслуживают препараты пептидной природы. Несмотря на их разнообразие, их объединяет ряд общих характеристик: низкая дозировка, отсутствие выраженных токсических эффектов, мягкость и длительность воздействия.
Одним из примеров успешной разработки, основанной на перечисленных выше принципах, является Кортексин — препарат, эффективность которого доказана на всех возможных уровнях исследования: клиническом, биологическом, клеточном, генетическом и молекулярном.
По данным МРТ в правой височной области головного мозга определяется очаг поражения, объем которого отчетливо нарастает к 3 суткам. При таком поражении на 28 сутки обычно наблюдается формирование глиального рубца и постинсультных кист. При применении Кортексина, когда пациент с ишемическим инсультом начинает получать препарат с первых часов заболевания, наряду с заметным улучшением общего самочувствия, клинической и неврологической картины, объем очага поражения мозга к 28 суткам уменьшается на 40%.
Терминология: Ишемия — Недостаточное кровоснабжение какого-либо органа или участка ткани, вызванное закупоркой или сужением соответствующей артерии; АТФ — Аденозинтрифосфат — нуклеотид, играет исключительно важную роль в обмене энергии и веществ в организмах; в первую очередь соединение известно как универсальный источник энергии для всех биохимических процессов, протекающих в живых системах. Деполяризация клеточной мембраны

Нейропротекторное противоапоптозное действие
Кортексин® является нейропротектором, который обладает терапевтическим воздействием, начиная с первых часов после ишемического поражения мозга. Это означает, что основной его мишенью является зона пенумбры — участок нервной ткани, окружающей очаг поражения, испытывающей кислородное и энергетическое голодание, но временно, до 6 часов, остающейся живой. От исхода этого процесса зависит возможность последующего восстановления нервных функций, жизнь и смерть больного. Кортексин
Глутамат является основным возбуждающим нейромедиатором нервной системы. При инсульте происходит избыточное высвобождение глутамата, приводящее к запуску каскада процессов, лежащих в основе гибели нейронов.
Восстановление синтеза АТФ
Аденозинтрифосфат (сокр. АТФ) — нуклеотид, играющий исключительно важную роль в обмене энергии и веществ в организмах, универсальный источник энергии для всех клеток организма. Падение содержания АТФ в клетках мозга является центральным звеном всех патологических процессов, протекающих на фоне ишемии мозга. Снижение синтеза и увеличение расхода АТФ показано сразу после начала ишемизации нервной ткани (Сорокина и др. , 2007). Недавние исследования продемонстрировали, что Кортексин
, 2007). Недавние исследования продемонстрировали, что Кортексин
Исследование продемонстрировало способность Кортексина® запускать процессы естественного восстановления АТФ в митохондриях нервных клеток. Поскольку падение уровня АТФ является одной из основных причин, приводящих к гибели нервных клеток при инсульте, восстановление этого показателя под действием Кортексина® объясняет его клиническую эффективность (Гранстрем О.К. и др., 2008).
Подавление отсроченной кальциевой дисрегуляции (ОКД)
При ишемии мозга и инсульте происходит активное проникновение ионов кальция в нейроны, что приводит к необратимому повышению их концентрации в клетке и последующему нарушению функционирования митохондрий, сопряженным с падением митохондриального потенциала (ΔΨm) (Ходоров и др., 2001; Krieger C. & Duchen M.R., 2002). Как правило, клетки, в которых происходит коллапс ΔΨm, после отмены глутамата не восстанавливают исходный потенциал и, в конечном итоге, погибают — наступает так называемая  , 1997; Сорокина Е. Г. и др., 2007).
, 1997; Сорокина Е. Г. и др., 2007).
Исследования митохондрильного потенциала (ΔΨm) методом флуоресцентной микроскопиидемонстрируют, что Кортексин значительно замедляет развитие отсроченной кальциевой дисрегуляции при действии глутамата. Представленная на рисунке запись митохондриальных потенциалов нейронов свидетельствует о сберегающем, защитном действии Кортексина
Нейротрофическое действие
Пептиды Кортексина® оказывают прямое и опосредованное нейротрофическое воздействие на клетки. Основные механизмы этого влияния базируются на изменении работы генов, регулирующих синтез собственных нейротрофических факторов таких, как мозговой нейротрофический фактор (BDNF) и фактор роста нервов (NGF).
Стимуляция роста нейритов в культуре головного мозга эмбриона цыпленка. В культуренервной ткани рост нейритов (отростоков нервной клетки, по которому нервные импульсы идут от тела клетки к органам и другим нервным клеткам) происходит только в присутствии нейротрофических факторов. В этом тесте проба с Кортексином® позволяет определить степень его нейротрофического воздействия: на правой микро-фотографии все поле вокруг островка нервной ткани занято развлетвленной сетью нейритов, в то время как, в контроле (левая микро-фотография) рост нейрональных отростков практически не наблюдается (на фотографиях приведены результы тестирования серии препарата. Подобное тестирование регулярно осуществляется в аналитической лаборатории научно-исследовательского центра ГК «ГЕРОФАРМ».
Таким образом, многочисленные независимые исследования убедительно демонстрируют наличие у Кортексина® множественных эффектов, затрагивающих каскадную регуляцию апоптоза, экспрессию нейтрофических факторов, энергетическое обеспечение нервной клетки и митохондриальный потенциал, функционирование рецепторов глутамата и регулирование концентрации ионов кальция в клетке, что в комплексе обеспечивает нейропротекторное и нейротрофическое действие препарата, а, в итоге, высокую эффективность лечения и улучшение качества жизни пациента.
Конкретные результаты клинического опыта отечественной медицины применения Кортексина® более подробно отражены в разделе Научные публикации
Литература:
- Герасимова М. М., Петушков А. Ю. / Влияние Кортексина на цитокиновый обмен при пояснично-крестцовых радикулопатиях. // Нейроиммунология. — 2004. — том II. — № 2. — С. 26.
- Гранстрем О.К., Сорокина Е.Г., Сторожевых Т.П., Штучная Г.В., Пинелис В.Г., Дьяконов М.М. / Последние новости о Кортексине (нейропротекция на молекулярном уровне). // Terra Medica Nova. — №5. — 2008. — С. 40-44.
- Королева С. В., Ашмарин И. П. / Разработка и применение экспертной системы анализа функционального континуума регуляторных пептидов» // Биоорганическая химия. — 2006. — Т. 32. — № 3 — С. 249–257.
- Скоромец А. А., Стаховская Л. В., Белкин А. А., Шеховцова К. В., Кербиков О. Б., Буренчев Д. В., Гаврилова О. В., Скворцова В. И. / Новые возможности нейропротекции в лечении ишемического инсульта // Журнал неврологии и психиатрии имени С.
 С. Корсакова. 2008. — Т. 22. — С.32–38.
С. Корсакова. 2008. — Т. 22. — С.32–38. - Сорокина Е. Г., Реутов В. П., Сенилова Я. Е., Ходоров Б.И., Пинелис В. Г. / Изменение содержания АТФ в зернистых клетках мозжечка при гиперстимуляции глутаматных рецепторов: возможное участие NO и нитритных ионов // Бюлл. эксперим. биол. и мед. — 2007. — № 4. — С. 419- 422.
- Ходоров Б.И., Сторожевых Т. П., Сурин А. М., Сорокина Е. Г., Юравичус А. И., Бородин А. В., Винская Н. П., Хаспеков Л. Г., Пинелис В. Г. / Митохондриальная деполяризация играет доминирующую роль в механизме нарушения нейронального кальциевого гомеостаза, вызванного глутаматом // Биол. мембраны. — 2001. — Т. 18, N 6. — С. 421–432.
- De Wied D. / Neuropeptides in learning and memory processes. // Behav. Brain. Res. — 1997. — Vol. 83. — P. 83–90.
- Krieger C. and Duchen MR. / Mitochondria, Ca2+ and neurodegenerative disease. // Eur. J. Pharmacol. — 2002. — Vol. 447. — P. 177–188.
- O’Collins VE., Macleod MR. , Donnan GA., Horky LL.,. van der Worp BH, and Howells DW. «1,026 Experimental Treatments in Acute Stroke» // Annals of Neurology. — 2006. — 59:467–477.
- Pinelis V. G., Storozhevykh T. P., Surin A. M., Senilova Ya.E., Persiyantzeva N. F., Tukhmatova G. R., Andreeva L. A., Myasoedov N. F., Granstrem O. «Neuroprotective effects of cortagen, cortexin and semax on glutamate neurotoxicity» / 30th European Peptide Symposium (30EPS), Helsinki, 30 August — 5 September 2008.
 С. Корсакова. 2008. — Т. 22. — С.32–38.
С. Корсакова. 2008. — Т. 22. — С.32–38. , Donnan GA., Horky LL.,. van der Worp BH, and Howells DW. «1,026 Experimental Treatments in Acute Stroke» // Annals of Neurology. — 2006. — 59:467–477.
, Donnan GA., Horky LL.,. van der Worp BH, and Howells DW. «1,026 Experimental Treatments in Acute Stroke» // Annals of Neurology. — 2006. — 59:467–477.как правильно и чем разводить
Как использовать
Перед тем, как делать инъекцию, нужно решить, с каким препаратом Кортексин будет разведен. Самый оптимальный вариант – проконсультироваться с медиком. Взрослым вводить можно любые препараты при условии отсутствия индивидуальной непереносимости лекарств, а вот в случае с детьми нужно быть предельно осторожными, поскольку неправильно подобранное средство может спровоцировать сильнейший приступ аллергической реакции. Поэтому перед тем как развести медикамент нужно убедиться в том, что жидкость безопасна для ребенка. Как правило, порошок разводят такими препаратами:
Как правило, порошок разводят такими препаратами:
- стерильная вода для уколов;
- Новокаин;
- физиологический раствор.
Разводить препарат Кортексин Новокаином для детей не рекомендовано поскольку могут возникнуть побочные эффекты. Для малышей самый оптимальный вариант – это инъекционная вода, она полностью безопасна.
Для своего же удобства лекарство лучше покупать во флаконах по 2 или 5 мл. Так вы сможете весь раствор использовать за один раз, не придется ничего утилизировать.
Кортексин разводить нужно только в стерильных условиях, вскрывать флакончик запрещено.
Раствор готовим следующим образом:
- необходимо взять шприц и набрать в него выбранный растворитель;
- иглой проткнуть пробку на флаконе с Кортексином, туда выпустить жидкость;
- немного подождать, пока порошок полностью растворится, это важный момент, спешить не стоит;
- набрать однородную жидкость в шприц.

Узнать конкретную информацию о дозировке лучше всего у доктора.
Как выбрать растворитель
Прокаин
Разводить данным средством будет целесообразно, если требуется легкая анестезия. Важно помнить о том, что медикамент не рекомендовано колоть лицам, у которых имеется почечная, сердечная, печеночная недостаточность. Если вы собираетесь делать укол ребенку– так же этот растворитель лучше не использовать ввиду того, что могут возникнуть нежелательные побочные эффекты.
Новокаин
Более популярен, нежели первый препарат, но он также не рекомендован, если нужно сделать укол Кортексина ребенку. Обладает мягким анестезирующим эффектом, имеет свойство скапливаться в нервных волокнах, за счет чего и можно добиться снижения болевого импульса. Разводить Новокаином нельзя более 10 дней, но специалисты именно на такой промежуток времени и назначают инъекции Кортексина.
Хлорид натрия
Важно купить средство в ампулах – иногда он выпускается в больших объемах для ингаляций, или промывки носа. Для взрослых людей максимальная доза препарата составляет 300 мл в день. О том, в каком количестве разводить средство для укола, лучше проконсультироваться с лечащим врачом, поскольку дозировка для взрослых и детей варьируется, он сможет назначить правильный объем вещества.
Для взрослых людей максимальная доза препарата составляет 300 мл в день. О том, в каком количестве разводить средство для укола, лучше проконсультироваться с лечащим врачом, поскольку дозировка для взрослых и детей варьируется, он сможет назначить правильный объем вещества.
Вода для инъекций
Ставить уколы Кортексин лучше именно с применением этого растворителя, он самый безопасный, подходит как для детей, так и для взрослых, не имеет противопоказаний. Важно: обычная и инъекционная вода – абсолютно две разных вещи! Для уколов жидкость нужно покупать в аптеке, она стерильна и продается в специальных ампулах. Разводить Кортексин с водой для инъекций нужно в четких пропорциях – о них подробно пациенту расскажет доктор.
Лидокаин
Прежде, чем развести лидокаином Кортексин, важно посоветоваться со специалистом, поскольку данный препарат не всегда рекомендуют использовать совместно с ноотропами. Аналог Новокаина, также имеет довольно мощный анестезирующий эффект. Однако стоит задуматься о целесообразности применении данного вещества. Кортексин – это ноотроп, колоть препараты этой группы препаратов не болезненно, то есть, особых показаний к использованию обезболивающих нет. Кроме того, в порошке Кортексина присутствуют аминокислоты, которые Лидокаин может разрушить, а значит, и терапия будет менее эффективной.
Однако стоит задуматься о целесообразности применении данного вещества. Кортексин – это ноотроп, колоть препараты этой группы препаратов не болезненно, то есть, особых показаний к использованию обезболивающих нет. Кроме того, в порошке Кортексина присутствуют аминокислоты, которые Лидокаин может разрушить, а значит, и терапия будет менее эффективной.
Источники:
Видаль: https://www.vidal.ru/drugs/cortexin__21252
ГРЛС: https://grls.rosminzdrav.ru/Grls_View_v2.aspx?routingGuid=5b83c52a-a651-44d6-8a17-429a3b239fee&t=
Нашли ошибку? Выделите ее и нажмите Ctrl + Enter
Кортексин: инструкция, цена, аналоги | лиофилизат для раствора для инъекций Герофарм
фармакодинамика. Кортексин — комплекс водорастворимых полипептидных фракций с молекулярной массой не более 10 000 Да.
Кортексин содержит комплекс низкомолекулярных водорастворимых полипептидных фракций, проникающих через ГЭБ непосредственно к нервным клеткам. Препарат оказывает ноотропное, нейропротекторное, антиоксидантное и тканеспецифическое действие.
Препарат оказывает ноотропное, нейропротекторное, антиоксидантное и тканеспецифическое действие.
Ноотропное — улучшает функции коры головного мозга, процессы обучения и памяти, концентрацию внимания, устойчивость при различных стрессовых воздействиях. Нейропротекторное — защищает нейроны от поражения различными эндогенными нейротоксическими факторами (глутамат, ионы кальция, свободные радикалы), снижает токсические эффекты психотропных веществ.
Антиоксидантное — ингибирует ПОЛ в нейронах, повышает выживаемость нейронов в условиях оксидативного стресса и гипоксии.
Тканеспецифическое — активирует метаболизм нейронов ЦНС и периферической нервной системы, репаративные процессы, способствует улучшению функций коры головного мозга и общего тонуса нервной системы.
Механизм действия препарата Кортексин обусловлен активацией пептидов нейронов и нейротрофических факторов мозга, оптимизацией баланса метаболизма возбуждающих и тормозных аминокислот, дофамина, серотонина, ГАМК-ергическим воздействием, снижением уровня пароксизмальной судорожной активности мозга, способностью улучшать его биоэлектрическую активность, предотвращением образования свободных радикалов (продуктов ПОЛ).
Фармакокинетика. Состав препарата Кортексин, действующим веществом которого является комплекс полипептидных фракций, не позволяет провести обычный фармакокинетический анализ отдельных компонентов.
в составе комплексной терапии черепно-мозговых травм, нарушений мозгового кровообращения, астенических состояний, энцефалопатии различного генеза, острых и хронических энцефалитов и энцефаломиелитов, эпилепсии, нарушений памяти и мышления, снижения способности к обучению, различных форм детского церебрального паралича, задержки психомоторного и речевого развития у детей.
препарат вводят в/м.
Содержимое флакона перед инъекцией растворяют в 1–2 мл воды для инъекций или 0,9% р-ра натрия хлорида, направляя иглу к стенке флакона во избежание пенообразования. Вводят ежедневно 1 раз в сутки: взрослым — в дозе 10 мг в течение 10 дней; детям с первых дней жизни с массой тела до 20 кг — в дозе 0,5 мг/кг массы тела; детям с массой тела >20 кг — в дозе 10 мг на протяжении 10 дней.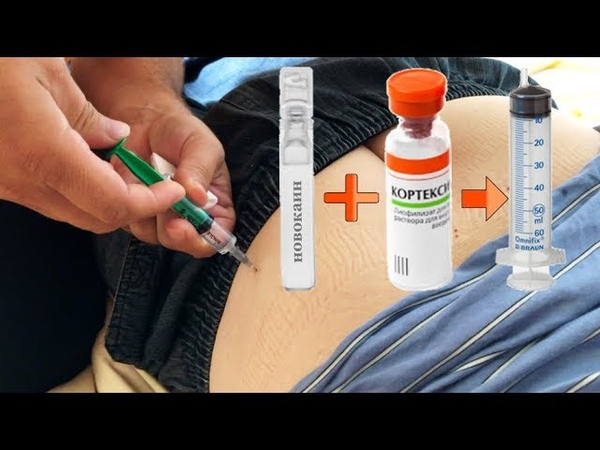
При необходимости проводят повторный курс лечения через 3–6 мес.
У взрослых целесообразно использовать Кортексин лиофилизат для р-ра для инъекций 10 мг.
При полушарном ишемическом инсульте в острый и ранний восстановительный период в дозе 10 мг 2 раза в сутки (утром и днем) в течение 10 дней с повторным курсом через 10 дней.
индивидуальная повышенная чувствительность к препарату.
в единичных случаях возможны реакции гиперчувствительности, в том числе гиперемия, зуд, сыпь на коже; тошнота.
Возможны изменения в месте введения: гиперемия, зуд, отек, которые самостоятельно исчезают в течение непродолжительного времени.
применять Кортексин только по назначению врача!
Не рекомендуется использовать 0,5% р-р прокаина для приготовления р-ра препарата.
Флакон с растворенным лекарственным препаратом нельзя хранить и использовать после хранения. Р-р препарата Кортексин не рекомендуется смешивать с другими р-рами.
Особенности действия лекарственного препарата при первом приеме или при его отмене отсутствуют.
В случае пропуска инъекции не рекомендуется вводить двойную дозу, а провести следующую инъекцию как обычно, в определенный день.
Специальных мер предосторожности при уничтожении неиспользованного лекарственного препарата не требуется.
Применение в период беременности и кормления грудью. Применение препарата в период беременности противопоказано. При необходимости назначения препарата кормление грудью следует прекратить.
Дети. Препарат можно применять в педиатрической практике.
Способность влиять на скорость реакции при управлении транспортными средствами или работе с другими механизмами. Не влияет.
несовместим с другими препаратами пептидной структуры.
Несовместимость. Не рекомендуется вводить в одном шприце с другими лекарственными средствами. Не рекомендуют использовать в качестве растворителя новокаин.
случаи передозировки не описаны.
в сухом, защищенном от света месте при температуре не выше 25 °С.
Дата добавления: 17.11.2021 г.
выполнение внутримышечной инъекции (в/м инъекции)
Врач прописал вам лекарство, которое должно вводиться внутримышечно (в/м инъекция; intramuscular or IM injection). При в/м инъекции лекарство вводится в крупные мышцы тела посредством шприца и иглы. В/м инъекции обычно делают в ягодицы, бедро, верхнюю часть бедра или плечо.
В больнице вам показывали, как делать в/м инъекции. Эта инструкция поможет вам вспомнить всю процедуру, когда вы будете выполнять ее дома.
Название вашего лекарства (medication name):
________________________
Доза вашего лекарства (amount per injection):
________________________
Принимайте Ваше лекарство каждые ________ часа (times per day)
Домашний уход
Подготовьте ваш комплект:
лекарство
шприц
2 иглы (одна для наполнения шприца лекарством из бутылочки, вторая для выполнения инъекции)
спиртовые тампоны
Емкость с твердыми непротыкаемыми стенками для утилизации использованных игл и шприцов (подойдет большая стеклянная банка с крышкой)
Тщательно вымойте руки до и после всех в/м инъекций.
Подготовьте лекарство:
Убедитесь, что вы взяли верное лекарство. Прочтите информацию на этикетке бутылочки, убедитесь, что наименование совпадает с прописанным вам лекарством.
Проверьте срок годности. Если срок лекарства истек, выбросьте его.
Не используйте лекарство, изменившее свой цвет, или в котором имеются посторонние включения.
Каждый раз протирайте пробку бутылочки спиртовым тампоном. Не прикасайтесь к пробке бутылочки, после того как вы протерли ее спиртом.
Подготовьте иглу и шприц:
Присоедините иглу к шприцу, не снимая колпачка с иглы.
Снимите колпачок с иглы и потяните за верхнюю часть шприца (поршень).
Наберите в шприц воздух в количестве, равном количеству лекарства, которое нужно принять. Будьте осторожны. Не прикасайтесь к игле.
Введите иглу в бутылочку через резиновую пробку на бутылочке и надавите на поршень, чтобы ввести воздух в бутылочку.
Оставьте шприц в бутылочке. Теперь переверните бутылочку вверх дном так, чтобы бутылочка была сверху, а шприц снизу.
Потяните за поршень. Благодаря этому лекарство будет поступать в шприц.
Когда будет набрано нужное количество лекарства, прекратите тянуть поршень вниз, однако иглу из бутылочки не извлекайте.
Проверьте, есть ли в шприце пузырьки воздуха. Чтобы удалить пузырьки воздуха:
Постучите по корпусу шприца кончиками или фалангами пальцев, чтобы пузырьки поднялись в верхнюю часть шприца.
Нажмите на поршень, чтобы выдавить пузырьки воздуха из шприца.
Еще раз потяните поршень вниз, чтобы набрать в шприц лекарство.
Повторяйте эти действия, пока не выйдут все пузырьки воздуха.
Если ваш врач сказал вам, что надо заменить иглу на новую, тогда поступайте так, как вам рекомендовали.
Извлеките иглу из бутылочки.
Если ваш врач сказал вам, что надо заменить иглу на новую, тогда поступайте так, как вам рекомендовали.
Выберите место укола.
Выбирайте места вдали от основных кровеносных сосудов и нервов. В больнице вам показывали эти участки тела.
Меняйте место укола для каждой инъекции. Избегайте постоянного использования одного и того же места укола.
Избегайте участков с гематомами, шрамами, опухолью, а также болезненных участков.
Подготовьтесь к введению лекарства:
Протрите место укола спиртом, и прежде чем делать укол, подождите, пока оно высохнет.
Держите шприц как дротик. Другой рукой растяните кожу вокруг места укола, не прикасаясь непосредственно к месту укола.
Введите лекарство:
Введите иглу под углом 90 градусов к поверхности подготовленного места укола быстрым движением, направленным вниз.
Отпустите кожу.
Придерживайте шприц, чтобы он не двигался.
Потяните за поршень.
Проверьте, есть ли в шприце кровь.
Если в шприце вы видите кровь, извлеките иглу, выбросьте шприц и начните все с начала с новым шприцем. Кровь в шприце означает, что вы попали в кровеносный сосуд. Никогда не вводите лекарство в кровеносный сосуд.
Если крови в шприце нет, нажимайте на поршень с постоянной скоростью. Введите все лекарство в шприце.
Извлеките иглу, вытянув шприц и иглу из тела движением вверх.
Приложите спиртовой тампон к месту укола.
Плотно прижмите тампон на одну минуту.
Осмотрите место укола на наличие красноты, кровотечения или гематомы.
При необходимости наложите на место укола повязку.
Никогда не надевайте колпачок на использованную иглу.
Выбросьте иглу и шприц в емкость с непротыкаемыми стенками.
Когда емкость заполнится, отнесите ее в ваше медицинское учреждение для должной утилизации биологически опасных отходов.
Прочие использованные материалы выбросьте в мусор.
Еще раз вымойте руки.




Последующее наблюдение
Запишитесь на прием к врачу в соответствии с рекомендациями нашего персонала.
Немедленно вызовите врача, если у вас наблюдаются какие-либо из следующих симптомов:
Игла сломалась и осталась в месте укола
Нет возможности самостоятельно сделать инъекцию
Непрекращающееся кровотечение в месте укола
Сильная боль в месте укола
Лекарство введено в неверный участок тела
Сыпь или опухоль в месте укола
Одышка
Температура тела выше 38,3°C (101°F)
Уколы кортексина — Вопрос невропатологу
Если вы не нашли нужной информации среди ответов на этот вопрос, или же ваша проблема немного отличается от представленной, попробуйте задать дополнительный вопрос врачу на этой же странице, если он будет по теме основного вопроса. Вы также можете задать новый вопрос, и через некоторое время наши врачи на него ответят. Это бесплатно. Также можете поискать нужную информацию в похожих вопросах на этой странице или через страницу поиска по сайту. Мы будем очень благодарны, если Вы порекомендуете нас своим друзьям в социальных сетях.
Это бесплатно. Также можете поискать нужную информацию в похожих вопросах на этой странице или через страницу поиска по сайту. Мы будем очень благодарны, если Вы порекомендуете нас своим друзьям в социальных сетях.Медпортал 03online.com осуществляет медконсультации в режиме переписки с врачами на сайте. Здесь вы получаете ответы от реальных практикующих специалистов в своей области. В настоящий момент на сайте можно получить консультацию по 74 направлениям: специалиста COVID-19, аллерголога, анестезиолога-реаниматолога, венеролога, гастроэнтеролога, гематолога, генетика, гепатолога, гериатра, гинеколога, гинеколога-эндокринолога, гомеопата, дерматолога, детского гастроэнтеролога, детского гинеколога, детского дерматолога, детского инфекциониста, детского кардиолога, детского лора, детского невролога, детского нефролога, детского онколога, детского офтальмолога, детского психолога, детского пульмонолога, детского ревматолога, детского уролога, детского хирурга, детского эндокринолога, дефектолога, диетолога, иммунолога, инфекциониста, кардиолога, клинического психолога, косметолога, липидолога, логопеда, лора, маммолога, медицинского юриста, нарколога, невропатолога, нейрохирурга, неонатолога, нефролога, нутрициолога, онколога, онкоуролога, ортопеда-травматолога, офтальмолога, паразитолога, педиатра, пластического хирурга, подолога, проктолога, психиатра, психолога, пульмонолога, ревматолога, рентгенолога, репродуктолога, сексолога-андролога, стоматолога, трихолога, уролога, фармацевта, физиотерапевта, фитотерапевта, флеболога, фтизиатра, хирурга, эндокринолога.
Мы отвечаем на 97.43% вопросов.
Оставайтесь с нами и будьте здоровы!
границ | Помимо перцептивной нагрузки и разбавления: обзор роли рабочей памяти в избирательном внимании
Введение
Вопрос о том, когда происходит визуальный отбор во время обработки информации, был основным вопросом в исследованиях избирательного внимания, который долгое время оставался нерешенным, принимая во внимание как ранний, так и поздний отбор. О раннем отборе свидетельствует обнаружение, среди прочего, отсутствия признаков идентификации неконтролируемой информации (например, Lachter et al., 2004). И наоборот, периферийные нерелевантные буквы-дистракторы, как правило, мешают идентификации центральных букв-мишеней, что позволяет предположить, что дистракторы были обработаны, по крайней мере, до уровня идентичности букв (например, Eriksen and Eriksen, 1974). Это означает поздний отбор. Теория перцептивной нагрузки (Lavie and Tsal, 1994; Lavie, 1995) предложила решение спора, предполагая, что локус отбора зависит от требований перцептивной обработки выполняемой задачи: высокие перцептивные требования предотвратят появление любой отвлекающей информации. обрабатываться, что приводит к раннему отбору, тогда как низкие требования к восприятию позволяют обрабатывать отвлекающие факторы, что требует позднего отбора.
обрабатываться, что приводит к раннему отбору, тогда как низкие требования к восприятию позволяют обрабатывать отвлекающие факторы, что требует позднего отбора.
Исследования, непосредственно изучающие эффект перцептивной нагрузки, долгое время, казалось, поддерживали идею о том, что релевантные требования перцептивной обработки определяют степень обработки нерелевантной информации (например, Lavie, 1995; Lavie and Cox, 1997; Lavie et al., 2004). Однако недавно было предложено альтернативное объяснение, предполагающее, что снижение отвлекаемости при высокой перцептивной нагрузке происходит из-за большего разбавления дистрактора (Benoni and Tsal, 2010, 2012; Tsal and Benoni, 2010; Wilson et al., 2011). С этой точки зрения, условия высокой перцептивной нагрузки связаны с пониженной отвлекаемостью просто потому, что отвлекающие факторы конкурируют с дополнительными релевантными не-целями в дисплеях с высокой нагрузкой, а не с истощением способности внимания при высокой перцептивной нагрузке.
Основное различие между моделями нагрузки и разбавления касается механизмов, лежащих в основе избирательности восприятия, тогда как обе модели предполагают, что дистракторы с большей вероятностью будут обрабатываться при определенных условиях восприятия.Тем не менее в таких случаях относительно экстенсивной обработки отвлекающих факторов поведение в значительной степени остается соответствующим цели. Другими словами, хотя обработка воспринимаемых отвлекающих факторов при низкой перцептивной нагрузке или низком разбавлении оказывает измеримое влияние на обработку цели, наблюдатели по-прежнему могут расставлять приоритеты в обработке цели и предотвращать, например, неадекватную реакцию на отвлекающую информацию. Таким образом, ключевой вопрос заключается в следующем: как поддерживаются приоритеты обработки для достижения целенаправленного поведения, когда текущие аспекты восприятия не позволяют обеспечить достаточную избирательность внимания, а отвлекающие факторы, вероятно, получат значительную обработку?
Разбавление включает в себя процесс раннего отбора: получает ли дистрактор обработку или нет, зависит от перцептивных характеристик визуального дисплея. Таким образом, модель разбавления не делает конкретных прогнозов относительно судьбы отвлекающей информации, которая не была исключена из обработки с помощью разбавления. Цал и Бенони (2010) действительно предлагают интерпретацию теории нагрузки, предполагая, что любое дополнительное увеличение нагрузки на ресурсы внимания должно снизить вероятность того, что ресурсы внимания перейдут к отвлекающим факторам. С этой точки зрения увеличение нагрузки на внимание, даже если оно не влияет на перцептивные аспекты задачи, должно быть связано со снижением обработки отвлекающих факторов.Как мы указываем ниже, это не так, и увеличение нагрузки на внимание, которое включает в себя контроль внимания сверху вниз, а не процессы выбора, определяемые стимулом, такие как перцептивная нагрузка, имеет тенденцию иметь эффект усиления обработки отвлекающих факторов.
Таким образом, модель разбавления не делает конкретных прогнозов относительно судьбы отвлекающей информации, которая не была исключена из обработки с помощью разбавления. Цал и Бенони (2010) действительно предлагают интерпретацию теории нагрузки, предполагая, что любое дополнительное увеличение нагрузки на ресурсы внимания должно снизить вероятность того, что ресурсы внимания перейдут к отвлекающим факторам. С этой точки зрения увеличение нагрузки на внимание, даже если оно не влияет на перцептивные аспекты задачи, должно быть связано со снижением обработки отвлекающих факторов.Как мы указываем ниже, это не так, и увеличение нагрузки на внимание, которое включает в себя контроль внимания сверху вниз, а не процессы выбора, определяемые стимулом, такие как перцептивная нагрузка, имеет тенденцию иметь эффект усиления обработки отвлекающих факторов.
Теория нагрузки предполагает, что функции когнитивного контроля, поддерживаемые лобными долями, особенно рабочая память, имеют решающее значение при позднем отборе (Lavie et al. , 2004; Lavie, 2010). С этой точки зрения рабочая память играет ключевую роль в поддержании приоритетов обработки, так что информация, связанная с целью и отвлекающим фактором, остается четко разделенной при обработке, и поведение может быть успешно направлено на информацию, относящуюся к задаче.Эта идея не нова, и более раннее предположение о том, что рабочая память и избирательное внимание могут функционально перекрываться, появилось в контексте модели рабочей памяти Баддели (Baddeley, 1996), в которой утверждалось, что ключевой функцией центрального исполнительного компонента рабочей памяти является для облегчения избирательного внимания к соответствующей информации в присутствии потенциальных отвлекающих факторов. В то время доказательства связи между рабочей памятью и избирательным вниманием были косвенными и исходили из двух направлений исследований.Во-первых, исследования возрастных изменений избирательного внимания показали, что способность предотвращать отвлечение внимания на ненужную информацию непропорционально зависит от возраста (например, Rabbitt, 1965; Hasher and Zacks, 1988).
, 2004; Lavie, 2010). С этой точки зрения рабочая память играет ключевую роль в поддержании приоритетов обработки, так что информация, связанная с целью и отвлекающим фактором, остается четко разделенной при обработке, и поведение может быть успешно направлено на информацию, относящуюся к задаче.Эта идея не нова, и более раннее предположение о том, что рабочая память и избирательное внимание могут функционально перекрываться, появилось в контексте модели рабочей памяти Баддели (Baddeley, 1996), в которой утверждалось, что ключевой функцией центрального исполнительного компонента рабочей памяти является для облегчения избирательного внимания к соответствующей информации в присутствии потенциальных отвлекающих факторов. В то время доказательства связи между рабочей памятью и избирательным вниманием были косвенными и исходили из двух направлений исследований.Во-первых, исследования возрастных изменений избирательного внимания показали, что способность предотвращать отвлечение внимания на ненужную информацию непропорционально зависит от возраста (например, Rabbitt, 1965; Hasher and Zacks, 1988). Наряду с обнаружением того, что производительность рабочей памяти также ухудшается с возрастом (например, Welford, 1958; Morris et al., 1988), это дало косвенные доказательства того, что рабочая память может каким-то образом участвовать в избирательной обработке информации. Второе раннее предположение о том, что рабочая память может быть вовлечена в избирательное внимание, было сделано Дезимоном и Дунканом (1995), которые утверждали, что контроль внимания сверху вниз (т.т. е., не полностью основанный на привлекающих внимание свойствах ввода), включает в себя поддержание шаблона, определяющего, какая информация имеет отношение к поставленной задаче, функция, идеально подходящая для рабочей памяти.
Наряду с обнаружением того, что производительность рабочей памяти также ухудшается с возрастом (например, Welford, 1958; Morris et al., 1988), это дало косвенные доказательства того, что рабочая память может каким-то образом участвовать в избирательной обработке информации. Второе раннее предположение о том, что рабочая память может быть вовлечена в избирательное внимание, было сделано Дезимоном и Дунканом (1995), которые утверждали, что контроль внимания сверху вниз (т.т. е., не полностью основанный на привлекающих внимание свойствах ввода), включает в себя поддержание шаблона, определяющего, какая информация имеет отношение к поставленной задаче, функция, идеально подходящая для рабочей памяти.
В первом исследовании, в котором были представлены прямые доказательства роли рабочей памяти в избирательном внимании, использовалась парадигма, сочетающая задание на рабочую память с заданием на избирательное внимание для измерения вмешательства дистрактора в контексте различной нагрузки на рабочую память (De Fockert et al. , 2001). ).В этом исследовании испытуемые выполняли задачу классификации имени цели (поп-звезда, политик), игнорируя отвлекающие лица (Young et al., 1986), так что любая обработка нерелевантных лиц приводила к снижению производительности в испытаниях, в которых категория лица была выбрана. несовместимы с текущей целевой категорией имени (например, имя Элтона Джона, сопровождаемое лицом Билла Клинтона), по сравнению с испытаниями, в которых категории имени и лица были либо совместимыми (например, имя и лицо Элтона Джона), либо не связанными ( е.г., имя Элтон Джон с анонимным лицом). Задача избирательного внимания выполнялась в контексте либо низкой, либо высокой нагрузки на рабочую память. В начале каждого испытания испытуемые видели набор из пяти цифр, которые они должны были запомнить до конца испытания, чтобы иметь возможность ответить на тест памяти. Нагрузкой рабочей памяти манипулировали, изменяя порядок установленных цифр: в испытаниях с низкой нагрузкой цифры всегда появлялись в последовательном порядке, тогда как в испытаниях с высокой нагрузкой использовался другой случайный порядок.
, 2001). ).В этом исследовании испытуемые выполняли задачу классификации имени цели (поп-звезда, политик), игнорируя отвлекающие лица (Young et al., 1986), так что любая обработка нерелевантных лиц приводила к снижению производительности в испытаниях, в которых категория лица была выбрана. несовместимы с текущей целевой категорией имени (например, имя Элтона Джона, сопровождаемое лицом Билла Клинтона), по сравнению с испытаниями, в которых категории имени и лица были либо совместимыми (например, имя и лицо Элтона Джона), либо не связанными ( е.г., имя Элтон Джон с анонимным лицом). Задача избирательного внимания выполнялась в контексте либо низкой, либо высокой нагрузки на рабочую память. В начале каждого испытания испытуемые видели набор из пяти цифр, которые они должны были запомнить до конца испытания, чтобы иметь возможность ответить на тест памяти. Нагрузкой рабочей памяти манипулировали, изменяя порядок установленных цифр: в испытаниях с низкой нагрузкой цифры всегда появлялись в последовательном порядке, тогда как в испытаниях с высокой нагрузкой использовался другой случайный порядок. Прогноз заключался в том, что если рабочая память важна для поддержания избирательного внимания к релевантным именам целей, то сделать ее относительно недоступной для этого (путем вовлечения рабочей памяти в дополнительную задачу с высокой нагрузкой) должно привести к менее избирательной обработке. Результаты подтвердили это предсказание. Эффекты совместимости с точки зрения времени реакции и показателей точности были выше при высокой нагрузке на рабочую память (разница в 78 мс между совместимыми и несовместимыми дисплеями) по сравнению с низкой нагрузкой (разница в 46 мс).Кроме того, нейронный ответ в областях мозга, предназначенных для обработки нерелевантных лиц, также был выше при высокой (по сравнению с низкой) нагрузке на рабочую память. Следуя первоначальным косвенным предположениям о роли рабочей памяти в избирательном внимании (Desimone and Duncan, 1995; Baddeley, 1996), это дало первые убедительные доказательства того, что у одних и тех же участников и без изменения каких-либо свойств задачи на избирательное внимание работа можно показать, что память влияет на зрительную отвлекаемость.
Прогноз заключался в том, что если рабочая память важна для поддержания избирательного внимания к релевантным именам целей, то сделать ее относительно недоступной для этого (путем вовлечения рабочей памяти в дополнительную задачу с высокой нагрузкой) должно привести к менее избирательной обработке. Результаты подтвердили это предсказание. Эффекты совместимости с точки зрения времени реакции и показателей точности были выше при высокой нагрузке на рабочую память (разница в 78 мс между совместимыми и несовместимыми дисплеями) по сравнению с низкой нагрузкой (разница в 46 мс).Кроме того, нейронный ответ в областях мозга, предназначенных для обработки нерелевантных лиц, также был выше при высокой (по сравнению с низкой) нагрузке на рабочую память. Следуя первоначальным косвенным предположениям о роли рабочей памяти в избирательном внимании (Desimone and Duncan, 1995; Baddeley, 1996), это дало первые убедительные доказательства того, что у одних и тех же участников и без изменения каких-либо свойств задачи на избирательное внимание работа можно показать, что память влияет на зрительную отвлекаемость.
Первоначальная демонстрация роли рабочей памяти в избирательном внимании подтолкнула к дальнейшим исследованиям связи между этими конструкциями.Основная цель этой статьи — предоставить обзор имеющихся данных, сосредоточив внимание на вопросе о том, как доступность рабочей памяти влияет на отвлекаемость при избирательном внимании. Другие важные аспекты, касающиеся связи между рабочей памятью и избирательным вниманием, в том числе то, как внимание определяет, какая информация вводится в рабочую память и расставляется по приоритетам в ней (например, Oberauer, 2003; Gazzaley and Nobre, 2012), и как внимание может быть смещено в сторону содержание рабочей памяти (Awh and Jonides, 2001; Soto et al., 2008; Olivers et al., 2011), здесь не рассматриваются. Вместо этого мы сосредоточимся конкретно на исследованиях, которые манипулируют уровнем доступности рабочей памяти во время внимания, и измерим отвлекаемость как функцию либо нагрузки на рабочую память, либо объема рабочей памяти. Как мы увидим, большинство исследований действительно показывают, что недоступность рабочей памяти для избирательного внимания приводит к увеличению отвлекаемости, хотя мы также обсудим данные, свидетельствующие о том, что загрузка рабочей памяти может привести к снижению отвлекаемости при определенных обстоятельствах. Наконец, обсуждаются исследования, объясняющие механизм того, как рабочая память может влиять на избирательное внимание.
Наконец, обсуждаются исследования, объясняющие механизм того, как рабочая память может влиять на избирательное внимание.
Загрузка рабочей памяти и отвлекаемость
С момента первого экспериментального сообщения о том, что высокая нагрузка на рабочую память может привести к усилению обработки нерелевантной информации, в ряде исследований были обнаружены аналогичные эффекты. Связанное с нагрузкой усиление интерференционного эффекта от нерелевантных лиц в задаче поп-звезды/политика было воспроизведено в двух экспериментах (Pecchinenda and Heil, 2007, Experiments 1 и 2; но см. Jongen and Jonkman, 2011, пример, в котором работа нагрузка на память увеличивала отвлечение лица, но незначительно).В стандартной задаче фланкера типа Эриксена (Eriksen and Eriksen, 1974), в которой внимание должно быть выборочно направлено на целевую букву в присутствии отвлекающей фланкерной буквы, интерференционный эффект, производимый фланкером (измеряемый путем сравнения производительности с дисплеи, в которых дистрактор совместим или несовместим с целью) значительно усиливается при одновременном сохранении в памяти набора из шести цифр (по сравнению с одной цифрой) (Lavie et al. , 2004, Experiment 1; De Fockert et al., 2010; Ахмед и Де Фокерт, 2012b). Добавление задачи на рабочую память имеет аналогичный эффект увеличения обработки нерелевантных фланкеров, отличных от букв, таких как стрелки влево или вправо (Pratt et al., 2011). Как задача поп-звезды/политика, так и задачи фланкера производят интерференционные эффекты типа Струпа, при этом отвлекающие факторы (лица, фланкерные буквы или стрелки), которые следует игнорировать, релевантны задаче в той мере, в какой они связаны с возможной реакцией. Действительно, интерференция цветовых слов в классической задаче Струпа также больше, когда нагрузка на рабочую память высока (Stins et al., 2004). Дистракторы появляются при каждом испытании в этих задачах, что делает их присутствие совершенно предсказуемым. То, что высокая нагрузка на рабочую память увеличивает степень обработки таких отвлекающих факторов, предполагает, что рабочая память участвует в активном подавлении известных отвлекающих факторов.
, 2004, Experiment 1; De Fockert et al., 2010; Ахмед и Де Фокерт, 2012b). Добавление задачи на рабочую память имеет аналогичный эффект увеличения обработки нерелевантных фланкеров, отличных от букв, таких как стрелки влево или вправо (Pratt et al., 2011). Как задача поп-звезды/политика, так и задачи фланкера производят интерференционные эффекты типа Струпа, при этом отвлекающие факторы (лица, фланкерные буквы или стрелки), которые следует игнорировать, релевантны задаче в той мере, в какой они связаны с возможной реакцией. Действительно, интерференция цветовых слов в классической задаче Струпа также больше, когда нагрузка на рабочую память высока (Stins et al., 2004). Дистракторы появляются при каждом испытании в этих задачах, что делает их присутствие совершенно предсказуемым. То, что высокая нагрузка на рабочую память увеличивает степень обработки таких отвлекающих факторов, предполагает, что рабочая память участвует в активном подавлении известных отвлекающих факторов.
Аналогичный эффект нагрузки на рабочую память увеличения обработки дистракторов был показан для дистракторов, которые не имеют отношения к задаче и присутствие которых не вполне предсказуемо. В задачах визуального поиска наличие заметного одиночного дистрактора приводит к ухудшению производительности, которое обычно интерпретируется как отражение захвата внимания дистрактором (см. обзор Theeuwes, 2010).Например, поиск целевой формы происходит медленнее, когда один из нецелевых элементов отображения имеет уникальный цвет, даже если цвет не имеет значения на протяжении всего эксперимента (Theeuwes, 1992). Такие эффекты захвата внимания выше при высокой нагрузке на рабочую память (Boot et al., 2005; Lavie and De Fockert, 2005, 2006, но см. De Fockert and Theeuwes, 2012, пример, когда высокая нагрузка на рабочую память не модулирует эффекты захвата). ), предполагая, что рабочая память участвует в минимизации отвлечения внимания, вызванного одноэлементными цветами.Косвенно интерпретация того, что одноэлементный захват включает в себя когнитивный контроль, возможно, рабочую память, дополнительно подтверждается данными нейровизуализации, показывающими, что величина захвата в поведении отрицательно коррелирует с активностью в левой лобной коре (De Fockert et al.
В задачах визуального поиска наличие заметного одиночного дистрактора приводит к ухудшению производительности, которое обычно интерпретируется как отражение захвата внимания дистрактором (см. обзор Theeuwes, 2010).Например, поиск целевой формы происходит медленнее, когда один из нецелевых элементов отображения имеет уникальный цвет, даже если цвет не имеет значения на протяжении всего эксперимента (Theeuwes, 1992). Такие эффекты захвата внимания выше при высокой нагрузке на рабочую память (Boot et al., 2005; Lavie and De Fockert, 2005, 2006, но см. De Fockert and Theeuwes, 2012, пример, когда высокая нагрузка на рабочую память не модулирует эффекты захвата). ), предполагая, что рабочая память участвует в минимизации отвлечения внимания, вызванного одноэлементными цветами.Косвенно интерпретация того, что одноэлементный захват включает в себя когнитивный контроль, возможно, рабочую память, дополнительно подтверждается данными нейровизуализации, показывающими, что величина захвата в поведении отрицательно коррелирует с активностью в левой лобной коре (De Fockert et al. , 2004). В дополнение к активной минимизации эффекта захвата синглетонов рабочая память может также участвовать в простом обнаружении их присутствия. Активность в правой префронтальной коре коррелирует с величиной захвата поведения, на этот раз демонстрируя положительную связь и только при высокой одновременной нагрузке на рабочую память (De Fockert and Theeuwes, 2012).Это говорит о том, что рабочая память также может играть роль в обнаружении потенциального отвлечения: активность в правой префронтальной коре была выше у участников, которые испытывали относительно сильный захват внимания, но только тогда, когда нагрузка на рабочую память была высокой и присутствовал одиночный дистрактор.
, 2004). В дополнение к активной минимизации эффекта захвата синглетонов рабочая память может также участвовать в простом обнаружении их присутствия. Активность в правой префронтальной коре коррелирует с величиной захвата поведения, на этот раз демонстрируя положительную связь и только при высокой одновременной нагрузке на рабочую память (De Fockert and Theeuwes, 2012).Это говорит о том, что рабочая память также может играть роль в обнаружении потенциального отвлечения: активность в правой префронтальной коре была выше у участников, которые испытывали относительно сильный захват внимания, но только тогда, когда нагрузка на рабочую память была высокой и присутствовал одиночный дистрактор.
Другие результаты также подтверждают мнение о том, что нагрузка на рабочую память влияет на степень обработки отвлекающих факторов, даже если они не связаны напрямую с реакцией на задачу.Иллюзия Эббингауза (или Титченера) представляет собой визуальную иллюзию размера, при которой воспринимаемый размер целевого круга зависит от размера окружающих индукторных кругов, так что мишень, окруженная большими индукторами, имеет меньший воспринимаемый размер по сравнению с мишенью, окруженной индукторами. малые индукторы (например, Roberts et al., 2005). Величину иллюзии Эббингауза можно рассматривать как показатель избирательного внимания, поскольку более активная обработка нерелевантных для задачи индукторов должна привести к большей иллюзии.Действительно, люди, которые проявляют превосходное избирательное внимание при выполнении ряда задач, также очень слабо испытывают иллюзию Эббингауза (De Fockert et al., 2011; Caparos et al., 2012, 2013). Когда иллюзия Эббингауза измеряется во время выполнения одновременной задачи на рабочую память, наблюдатели испытывают большую иллюзию Эббингауза, когда они поддерживают большой набор цифр в рабочей памяти, по сравнению с небольшим набором (De Fockert and Wu, 2009). Было показано, что те же манипуляции с нагрузкой на рабочую память влияют на слепоту по невнимательности.Когда наблюдатели выполняют сложную перцептивную задачу, включающую сравнение размеров двух расположенных в центре линий, единичное появление неожиданного дополнительного визуального стимула часто остается незамеченным (Rock et al.
малые индукторы (например, Roberts et al., 2005). Величину иллюзии Эббингауза можно рассматривать как показатель избирательного внимания, поскольку более активная обработка нерелевантных для задачи индукторов должна привести к большей иллюзии.Действительно, люди, которые проявляют превосходное избирательное внимание при выполнении ряда задач, также очень слабо испытывают иллюзию Эббингауза (De Fockert et al., 2011; Caparos et al., 2012, 2013). Когда иллюзия Эббингауза измеряется во время выполнения одновременной задачи на рабочую память, наблюдатели испытывают большую иллюзию Эббингауза, когда они поддерживают большой набор цифр в рабочей памяти, по сравнению с небольшим набором (De Fockert and Wu, 2009). Было показано, что те же манипуляции с нагрузкой на рабочую память влияют на слепоту по невнимательности.Когда наблюдатели выполняют сложную перцептивную задачу, включающую сравнение размеров двух расположенных в центре линий, единичное появление неожиданного дополнительного визуального стимула часто остается незамеченным (Rock et al. , 1992; Mack and Rock, 1998). Однако такая слепота невнимания может быть устранена за счет высокой нагрузки на рабочую память (De Fockert and Bremner, 2011), что позволяет предположить, что критический стимул, не относящийся к задаче, с большей вероятностью будет восприниматься, когда рабочая память недоступна для удержания внимания на соответствующей задаче.Подобный эффект нагрузки рабочей памяти на обработку нерелевантного стимула, совершенно не относящегося к задаче, был показан Carmel et al. (2012), которые обнаружили, что нерелевантное лицо, представленное вместе с релевантной задачей категоризации имени, с большей вероятностью будет впоследствии идентифицировано, когда параллельная нагрузка на рабочую память была высокой. Интересно, что этот эффект нагрузки на рабочую память не был обнаружен, когда отвлекающими изображениями были здания, что позволяет предположить, что игнорирование значимых отвлекающих факторов (таких как лица) может быть особенно связано с наличием рабочей памяти.
, 1992; Mack and Rock, 1998). Однако такая слепота невнимания может быть устранена за счет высокой нагрузки на рабочую память (De Fockert and Bremner, 2011), что позволяет предположить, что критический стимул, не относящийся к задаче, с большей вероятностью будет восприниматься, когда рабочая память недоступна для удержания внимания на соответствующей задаче.Подобный эффект нагрузки рабочей памяти на обработку нерелевантного стимула, совершенно не относящегося к задаче, был показан Carmel et al. (2012), которые обнаружили, что нерелевантное лицо, представленное вместе с релевантной задачей категоризации имени, с большей вероятностью будет впоследствии идентифицировано, когда параллельная нагрузка на рабочую память была высокой. Интересно, что этот эффект нагрузки на рабочую память не был обнаружен, когда отвлекающими изображениями были здания, что позволяет предположить, что игнорирование значимых отвлекающих факторов (таких как лица) может быть особенно связано с наличием рабочей памяти.
Доказательства того, что рабочая память участвует в предотвращении обработки отвлекающих факторов, которые напрямую не связаны с какой-либо релевантной для задачи реакцией, важны, поскольку они предполагают, что влияние рабочей памяти на избирательное внимание происходит на уровне сенсорной обработки, а не реакции. выбор. Эффекты нагрузки рабочей памяти на захват внимания, иллюзию Эббингауза и обнаружение критического элемента при слепоте по невниманию вряд ли отражают более сильную тенденцию активировать неправильный ответный код после обработки отвлекающего фактора.Задача захвата внимания требует реагирования на ориентацию (горизонтальную, вертикальную) линии в целевой форме, а нецелевые формы никогда не содержат горизонтальных или вертикальных линий. Вместо этого отвлекающий синглтон определяется своим уникальным цветом, который не имеет отношения к задаче. В задаче Эббингауза профили реакции были бы противоположны наблюдаемым, если бы люди ошибочно реагировали на размер отвлекающих факторов. В задаче на слепоту по невнимательности критический элемент встречается только один раз и не связан ни с каким ответом на задачу.Таким образом, маловероятно, что рабочая память воздействует на избирательное внимание, предотвращая активацию реакций, связанных с отвлекающими факторами. Вместо этого увеличение при высокой нагрузке на рабочую память эффекта захвата внимания, иллюзии Эббингауза и частоты обнаружения при невнимательной слепоте, по-видимому, является результатом большего восприятия отвлекающих факторов при высокой нагрузке. Этот вывод дополнительно подтверждается данными фМРТ о том, что влияние рабочей памяти на обработку дистракторов можно наблюдать уже в V1 первичной зрительной коры (Kelley and Lavie, 2011).
В задаче на слепоту по невнимательности критический элемент встречается только один раз и не связан ни с каким ответом на задачу.Таким образом, маловероятно, что рабочая память воздействует на избирательное внимание, предотвращая активацию реакций, связанных с отвлекающими факторами. Вместо этого увеличение при высокой нагрузке на рабочую память эффекта захвата внимания, иллюзии Эббингауза и частоты обнаружения при невнимательной слепоте, по-видимому, является результатом большего восприятия отвлекающих факторов при высокой нагрузке. Этот вывод дополнительно подтверждается данными фМРТ о том, что влияние рабочей памяти на обработку дистракторов можно наблюдать уже в V1 первичной зрительной коры (Kelley and Lavie, 2011).
В дополнение к исследованиям, показывающим, что нагрузка на рабочую память приводит к большему вмешательству дистракторов, когда стимулы для обеих задач представлены визуально, есть также доказательства того, что обработка дистракторов увеличивается, когда задачи на рабочую память и избирательное внимание включают разные сенсорные модальности (Dalton et al. , 2009а,б). Высокая нагрузка на задание на рабочую память, включающее вокальное повторение визуально представленных цифр, во время задания, которое требовало внимания к целевым звукам и игнорирования отвлекающих звуков, приводила к большему вмешательству со стороны слуховых отвлекающих факторов (Dalton et al., 2009а). Тот же эффект увеличения вмешательства дистрактора при сохранении в рабочей памяти большого визуально представленного набора цифр был показан, когда внимание должно быть направлено на тактильную цель, игнорируя нерелевантный тактильный дистрактор (Dalton et al., 2009b). Однако, наоборот, разница в стоимости обработки зрительных целей, создаваемых либо тактильными, либо болевыми дистракторами, уменьшается после умеренного увеличения нагрузки на зрительную рабочую память (Legrain et al., 2011), результат, который можно интерпретировать как демонстрирующий, что работающая память привела к снижению эффекта захвата внимания болезненными отвлекающими факторами.Это говорит о том, что рабочая память в большей степени участвует в уменьшении помех от отвлекающих факторов в той же модальности, что и цель, по сравнению с теми, которые находятся в модальности, отличной от целевой модальности.
, 2009а,б). Высокая нагрузка на задание на рабочую память, включающее вокальное повторение визуально представленных цифр, во время задания, которое требовало внимания к целевым звукам и игнорирования отвлекающих звуков, приводила к большему вмешательству со стороны слуховых отвлекающих факторов (Dalton et al., 2009а). Тот же эффект увеличения вмешательства дистрактора при сохранении в рабочей памяти большого визуально представленного набора цифр был показан, когда внимание должно быть направлено на тактильную цель, игнорируя нерелевантный тактильный дистрактор (Dalton et al., 2009b). Однако, наоборот, разница в стоимости обработки зрительных целей, создаваемых либо тактильными, либо болевыми дистракторами, уменьшается после умеренного увеличения нагрузки на зрительную рабочую память (Legrain et al., 2011), результат, который можно интерпретировать как демонстрирующий, что работающая память привела к снижению эффекта захвата внимания болезненными отвлекающими факторами.Это говорит о том, что рабочая память в большей степени участвует в уменьшении помех от отвлекающих факторов в той же модальности, что и цель, по сравнению с теми, которые находятся в модальности, отличной от целевой модальности. Требуется дальнейшая работа, чтобы проверить это предположение, непосредственно сравнивая влияние дистракторов одинаковой (или другой) модальности на целевую обработку при различных уровнях нагрузки.
Требуется дальнейшая работа, чтобы проверить это предположение, непосредственно сравнивая влияние дистракторов одинаковой (или другой) модальности на целевую обработку при различных уровнях нагрузки.
Несмотря на то, что ряд экспериментальных задач и измерений в настоящее время свидетельствуют о том, что недоступность рабочей памяти во время избирательного внимания может увеличить обработку отвлекающих факторов, растущий объем работ, посвященных роли рабочей памяти в избирательном внимании, также иногда обнаруживал, что работающая память нагрузка на память может по-разному влиять на отвлекаемость.Неоднократно было показано, что высокая нагрузка на рабочую память либо увеличивает, либо снижает отвлекаемость в зависимости от того, перекрывается ли содержимое задачи рабочей памяти с обработкой цели или отвлекающего фактора в задаче избирательного внимания соответственно (Kim et al., 2005; Парк и др., 2007; де Лианьо и др., 2010). Высокая нагрузка на рабочую память, связанная с сохранением набора букв, приводит к большей обработке нерелевантного цвета слова цвета Струпа, когда необходимо учитывать значение слова (и игнорировать цвет), но к уменьшению обработки нерелевантного слова, когда цвет должен присутствовать (и значение слова игнорируется). Высокая нагрузка на задачу рабочей памяти для пространственной локации ни в том, ни в другом случае не влияет на работу дистрактора (Kim et al., 2005; de Liano et al., 2010). Точно так же, когда задание на рабочую память включает в себя запоминание либо лиц, либо домов, а задание на избирательное внимание также требует внимания к лицам и игнорирования домов, или наоборот, эффекты отвлекающих факторов усиливаются, когда элементы рабочей памяти относятся к той же категории, что и цели в задании. задача на внимание, но снижается, когда они совпадают с отвлекающими факторами (Park et al., 2007). Эти результаты подразумевают, что рабочая память может быть загружена для определенной категории стимулов и что загруженная категория должна перекрываться с целевой обработкой, чтобы привести к увеличению обработки отвлекающих факторов. Другая работа также предполагает, что когнитивная нагрузка может оказывать противоположное влияние на отвлекаемость в зависимости от характера отвлекающей информации.
Высокая нагрузка на задачу рабочей памяти для пространственной локации ни в том, ни в другом случае не влияет на работу дистрактора (Kim et al., 2005; de Liano et al., 2010). Точно так же, когда задание на рабочую память включает в себя запоминание либо лиц, либо домов, а задание на избирательное внимание также требует внимания к лицам и игнорирования домов, или наоборот, эффекты отвлекающих факторов усиливаются, когда элементы рабочей памяти относятся к той же категории, что и цели в задании. задача на внимание, но снижается, когда они совпадают с отвлекающими факторами (Park et al., 2007). Эти результаты подразумевают, что рабочая память может быть загружена для определенной категории стимулов и что загруженная категория должна перекрываться с целевой обработкой, чтобы привести к увеличению обработки отвлекающих факторов. Другая работа также предполагает, что когнитивная нагрузка может оказывать противоположное влияние на отвлекаемость в зависимости от характера отвлекающей информации. Бут и др. (2005) обнаружили, что в то время как привлечение внимания однотонными цветами увеличивалось при высокой когнитивной нагрузке, привлечение внимания внезапными приступами снижалось.Как показано ниже, значимость отвлекающего фактора может быть важным фактором, определяющим, повлияет ли нагрузка на рабочую память на обработку отвлекающих факторов.
Бут и др. (2005) обнаружили, что в то время как привлечение внимания однотонными цветами увеличивалось при высокой когнитивной нагрузке, привлечение внимания внезапными приступами снижалось.Как показано ниже, значимость отвлекающего фактора может быть важным фактором, определяющим, повлияет ли нагрузка на рабочую память на обработку отвлекающих факторов.
Как представление о том, что содержимое загрузки рабочей памяти должно перекрываться с обработкой цели (Kim et al., 2005; Park et al., 2007), согласуется с другими данными? Во многих исследованиях, показывающих увеличение отвлекаемости при высокой нагрузке на рабочую память, манипулирование нагрузкой включало сохранение наборов цифр или порядка цифр. Эта задача рабочей памяти действительно иногда перекрывалась с целевой обработкой задачи избирательного внимания.Когда задача избирательного внимания требовала обращать внимание на имена целей и игнорировать лица отвлекающих факторов (De Fockert et al., 2001), можно утверждать, что задача рабочей памяти имеет больше общего с вербальными требованиями классификации имен целей, чем с идентификацией отвлекающих факторов. лица. Другие результаты, однако, не очень хорошо согласуются с утверждением о том, что тип нагрузки на рабочую память должен демонстрировать большее совпадение с целью, чем с обработкой отвлекающих факторов, чтобы увеличить отвлекаемость. Наблюдается большая обработка дистрактора, когда задача рабочей памяти в равной степени перекрывается с обработкой цели и дистрактора, например, когда и цель, и дистрактор являются буквами (Lavie et al., 2004) или стрелки (Pratt et al., 2011). Более того, загрузка рабочей памяти цифрами также увеличивает нагрузку на отвлекающие факторы, когда целевая задача требует суждений о размере (De Fockert and Wu, 2009; De Fockert and Bremner, 2011), и даже когда задача избирательного внимания включает модальность, отличную от задачи рабочей памяти. (Далтон и др., 2009a,b). Кажется, что нагрузка на рабочую память способна увеличить отвлекаемость, даже если перекрытие между содержанием задачи на рабочую память и целевой обработкой при избирательном внимании минимально.
лица. Другие результаты, однако, не очень хорошо согласуются с утверждением о том, что тип нагрузки на рабочую память должен демонстрировать большее совпадение с целью, чем с обработкой отвлекающих факторов, чтобы увеличить отвлекаемость. Наблюдается большая обработка дистрактора, когда задача рабочей памяти в равной степени перекрывается с обработкой цели и дистрактора, например, когда и цель, и дистрактор являются буквами (Lavie et al., 2004) или стрелки (Pratt et al., 2011). Более того, загрузка рабочей памяти цифрами также увеличивает нагрузку на отвлекающие факторы, когда целевая задача требует суждений о размере (De Fockert and Wu, 2009; De Fockert and Bremner, 2011), и даже когда задача избирательного внимания включает модальность, отличную от задачи рабочей памяти. (Далтон и др., 2009a,b). Кажется, что нагрузка на рабочую память способна увеличить отвлекаемость, даже если перекрытие между содержанием задачи на рабочую память и целевой обработкой при избирательном внимании минимально.
В своей работе мы также сталкивались со случаями, когда высокая нагрузка на рабочую память не приводила к повышению уровня обработки дистракторов. В недавнем исследовании фМРТ мы обнаружили, что поведенческий эффект захвата внимания не усиливался при высокой нагрузке на рабочую память, даже несмотря на то, что реакция в нижней лобной извилине показала достоверную взаимосвязь между нагрузкой на рабочую память и наличием отвлекающего одиночного элемента (De Fockert and Тьювес, 2012). В этом исследовании были заблокированы условия, в которых присутствовал или отсутствовал цветовой синглтон.Возможно, это уменьшило заметность дистракторов, поскольку их присутствие было совершенно предсказуемо, в отличие от предыдущей работы, которая действительно показала влияние нагрузки на рабочую память на захват внимания цветными синглтонами, но в которых присутствовали тесты, в которых присутствовал дистрактор-синглетон. а отсутствовали либо случайным образом в пределах блока (Lavie and De Fockert, 2005), либо в которых одиночный цвет мог совпадать либо с мишенью, либо с дистрактором внутри блока (Boot et al. , 2005). Другая работа также показала, что нагрузка на рабочую память с меньшей вероятностью влияет на обработку ожидаемых отвлекающих факторов (Macdonald and Lavie, 2008).Кроме того, как показано Carmel et al. (2012), обработка дистрактора зависит от нагрузки рабочей памяти только тогда, когда дистрактор достаточно заметен (лица, а не здания в их исследовании), хотя влияние нагрузки зрительной рабочей памяти на интерференционные эффекты Саймона (вызванные нерелевантным расположением целевой стимул, возможно, особенно заметная форма вмешательства) менее ясна (Stins et al., 2004), а эмоциональные лица вызывают помехи независимо от уровня одновременной нагрузки на рабочую память (Pecchinenda and Heil, 2007, Experiment 3).В совокупности эти данные свидетельствуют о том, что заметность отвлекающих факторов является важным фактором, влияющим на то, влияет ли рабочая нагрузка на память на избирательное внимание. Влияние нагрузки рабочей памяти на интерференцию может иметь перевернутую U-образную функцию по мере увеличения заметности дистракторов: неброские дистракторы (например, здания) можно игнорировать даже при загруженной рабочей памяти, в то время как сильно выделяющиеся дистракторы (например, эмоциональные лица, нерелевантное целевое местоположение в задаче Саймона, внезапные синглтоны) обрабатываются даже при наличии рабочей памяти. Когда значимость отвлекающего фактора находится между этими крайними значениями (например, лица, буквы, связанные с ответом на задание, цветные синглтоны), рабочая память способна предотвратить обработку отвлекающих факторов. Необходима дальнейшая работа для систематического изучения роли отвлекающих факторов.
, 2005). Другая работа также показала, что нагрузка на рабочую память с меньшей вероятностью влияет на обработку ожидаемых отвлекающих факторов (Macdonald and Lavie, 2008).Кроме того, как показано Carmel et al. (2012), обработка дистрактора зависит от нагрузки рабочей памяти только тогда, когда дистрактор достаточно заметен (лица, а не здания в их исследовании), хотя влияние нагрузки зрительной рабочей памяти на интерференционные эффекты Саймона (вызванные нерелевантным расположением целевой стимул, возможно, особенно заметная форма вмешательства) менее ясна (Stins et al., 2004), а эмоциональные лица вызывают помехи независимо от уровня одновременной нагрузки на рабочую память (Pecchinenda and Heil, 2007, Experiment 3).В совокупности эти данные свидетельствуют о том, что заметность отвлекающих факторов является важным фактором, влияющим на то, влияет ли рабочая нагрузка на память на избирательное внимание. Влияние нагрузки рабочей памяти на интерференцию может иметь перевернутую U-образную функцию по мере увеличения заметности дистракторов: неброские дистракторы (например, здания) можно игнорировать даже при загруженной рабочей памяти, в то время как сильно выделяющиеся дистракторы (например, эмоциональные лица, нерелевантное целевое местоположение в задаче Саймона, внезапные синглтоны) обрабатываются даже при наличии рабочей памяти. Когда значимость отвлекающего фактора находится между этими крайними значениями (например, лица, буквы, связанные с ответом на задание, цветные синглтоны), рабочая память способна предотвратить обработку отвлекающих факторов. Необходима дальнейшая работа для систематического изучения роли отвлекающих факторов.
Отвлечение на девиантные звуки также снижается при высокой нагрузке на рабочую память (Berti and Schröger, 2003; SanMiguel et al., 2008), хотя увеличение нагрузки на рабочую память (от n = 0 до n = 1 в задаче n — назад) может иметь недостаточно загруженную рабочую память, что приводит к увеличению отвлекаемости в этих исследованиях.В другом недавнем исследовании мы обнаружили, что высокая нагрузка на рабочую память может привести к значительному снижению отвлекаемости в задаче Navon (Ahmed and De Fockert, 2012a). В задаче Навона глобальные стимулы состоят из множества локальных элементов, и внимание должно быть направлено либо на глобальную форму, либо на локальные элементы (Navon, 1977). Отвлекаемость может быть измерена путем сравнения производительности в условиях, в которых информация на посещаемом и необслуживаемом уровнях совместима (по сравнению с несовместимой). Когда внимание было направлено на локальный уровень, рабочая нагрузка памяти имела ожидаемый эффект увеличения отвлекаемости от глобального уровня.Однако, наоборот, отвлекающие эффекты уменьшались за счет высокой нагрузки на рабочую память, когда внимание было направлено на глобальный уровень, а локальный уровень приходилось игнорировать. Ниже мы обсудим эти результаты подробнее, когда будем рассматривать возможный механизм влияния нагрузки рабочей памяти на внимание.
Отвлекаемость может быть измерена путем сравнения производительности в условиях, в которых информация на посещаемом и необслуживаемом уровнях совместима (по сравнению с несовместимой). Когда внимание было направлено на локальный уровень, рабочая нагрузка памяти имела ожидаемый эффект увеличения отвлекаемости от глобального уровня.Однако, наоборот, отвлекающие эффекты уменьшались за счет высокой нагрузки на рабочую память, когда внимание было направлено на глобальный уровень, а локальный уровень приходилось игнорировать. Ниже мы обсудим эти результаты подробнее, когда будем рассматривать возможный механизм влияния нагрузки рабочей памяти на внимание.
Подводя итог работе с использованием манипуляций с нагрузкой на рабочую память во время избирательного внимания, можно сказать, что имеется много свидетельств того, что обработка информации, не относящейся к задаче, усиливается, когда нагрузка рабочей памяти на параллельную задачу высока, что подразумевает, что рабочая память играет роль в активный контроль против отвлекающих помех. Влияние рабочей памяти на избирательное внимание было продемонстрировано в задачах типа Струпа, где дистрактор связан с одним из ответов на задачу, а также в задачах, в которых дистрактор не может привести к активации ответа, что позволяет предположить, что эффект рабочей памяти имеет ранний локус внимания. Напротив, есть также примеры, в которых высокая рабочая нагрузка не привела к увеличению степени обработки дистракторов при избирательном внимании. К ним относятся ситуации, в которых содержание задачи на рабочую память в основном перекрывается с обработкой отвлекающего фактора (а не цели) при избирательном внимании (т.g., Kim et al., 2005), хотя есть много исследований, в которых высокая нагрузка на рабочую память действительно приводит к большей обработке отвлекающих факторов, несмотря на минимальное перекрытие между задачей на рабочую память и обработкой цели. Обработка отвлекающих факторов может с большей вероятностью увеличиваться при высокой нагрузке на рабочую память, когда отвлекающие факторы могут вызывать помехи либо потому, что они связаны с релевантной для задачи реакцией, либо потому, что они имеют промежуточный уровень значимости или возникают непредсказуемо от испытания к испытанию.
Влияние рабочей памяти на избирательное внимание было продемонстрировано в задачах типа Струпа, где дистрактор связан с одним из ответов на задачу, а также в задачах, в которых дистрактор не может привести к активации ответа, что позволяет предположить, что эффект рабочей памяти имеет ранний локус внимания. Напротив, есть также примеры, в которых высокая рабочая нагрузка не привела к увеличению степени обработки дистракторов при избирательном внимании. К ним относятся ситуации, в которых содержание задачи на рабочую память в основном перекрывается с обработкой отвлекающего фактора (а не цели) при избирательном внимании (т.g., Kim et al., 2005), хотя есть много исследований, в которых высокая нагрузка на рабочую память действительно приводит к большей обработке отвлекающих факторов, несмотря на минимальное перекрытие между задачей на рабочую память и обработкой цели. Обработка отвлекающих факторов может с большей вероятностью увеличиваться при высокой нагрузке на рабочую память, когда отвлекающие факторы могут вызывать помехи либо потому, что они связаны с релевантной для задачи реакцией, либо потому, что они имеют промежуточный уровень значимости или возникают непредсказуемо от испытания к испытанию.
В целом, доказательства большей обработки дистракторов при высокой нагрузке на рабочую память кажутся более убедительными, чем доказательства противоположного эффекта. Действительно, метаанализ величины эффекта, о котором сообщалось в 26 исследованиях (49 экспериментов) по манипулированию нагрузкой на рабочую память во время избирательного внимания, показывает, что преобладающим эффектом нагрузки на рабочую память является увеличение обработки дистракторов [среднее r = 0,202, t ( 48) = 2,95, СОС = 0,0686, р < 0,005; см. табл. 1].Это убедительный вывод, так как он включает в себя эксперименты, обнаружившие обратные эффекты (уменьшение отвлекаемости под нагрузкой), которые, тем не менее, были предсказаны на основе изменений пространственного профиля внимания, а также показали, что высокая нагрузка на рабочую память увеличивает обработку дистракторов, когда ожидается, что это произойдет (см. ниже; Ahmed and De Fockert, 2012a,b). Кроме того, значительно больше демонстраций повышения отвлекаемости при высокой нагрузке на рабочую память (35 экспериментов), чем снижения [14 экспериментов; χ 2 (1) = 9. 0, p < 0,01, двусторонний], хотя величина эффекта была одинаковой как в исследованиях, показывающих увеличение (среднее r = 0,477), так и в исследованиях, показывающих снижение (среднее r = 0,484, p). > 0,9). Мы отмечаем, что любая предвзятость в отношении публикации нулевых результатов означает, что, вероятно, существуют незарегистрированные неудачи в воспроизведении исходного эффекта, хотя такая предвзятость не повлияет на количество зарегистрированных достоверных обращений эффекта.
0, p < 0,01, двусторонний], хотя величина эффекта была одинаковой как в исследованиях, показывающих увеличение (среднее r = 0,477), так и в исследованиях, показывающих снижение (среднее r = 0,484, p). > 0,9). Мы отмечаем, что любая предвзятость в отношении публикации нулевых результатов означает, что, вероятно, существуют незарегистрированные неудачи в воспроизведении исходного эффекта, хотя такая предвзятость не повлияет на количество зарегистрированных достоверных обращений эффекта.
Таблица 1.Величина эффекта ( r ) для эффекта нагрузки рабочей памяти (WM) (если не указано иное) на обработку дистрактора (влияние времени реакции, если не указано иное) .
Вывод о том, что высокая нагрузка на рабочую память связана с повышенной обработкой отвлекающих факторов, согласуется с нагрузочной теорией избирательного внимания (Lavie et al., 2004). Согласно модели, активный контроль над обработкой воспринимаемых отвлекающих факторов требует наличия рабочей памяти, чтобы поддерживать четкое различие между релевантной и нерелевантной обработкой. Высокая нагрузка на рабочую память, которая должна выполняться одновременно с избирательным вниманием, оставляет меньше возможностей для определения приоритетов соответствующих целей, что приводит к большей отвлекаемости, обнаруженной в исследованиях, рассмотренных здесь.
Индивидуальные различия в объеме рабочей памяти и отвлекаемости
Обсуждаемые до сих пор доказательства роли рабочей памяти в обработке отвлекающих факторов были основаны на манипуляциях с нагрузкой на рабочую память, обычно у одних и тех же участников.Сходящиеся доказательства в пользу утверждения о том, что рабочая память влияет на степень обработки отвлекающих факторов при выборе, получены в исследованиях, изучающих избирательное внимание у людей с разным уровнем рабочей памяти. Обоснование заключается в том, что доступность рабочей памяти для избирательного внимания хронически снижена у людей с низким объемом рабочей памяти по сравнению с людьми с более высокими возможностями. Следовательно, различия в отвлекаемости между людьми с низким или высоким объемом рабочей памяти должны быть аналогичны различиям в отвлекаемости одного и того же человека с высокой или низкой нагрузкой на рабочую память соответственно.
Существует много свидетельств того, что эффективность внимания связана с объемом рабочей памяти, часто показывая, что люди с большим объемом рабочей памяти более эффективно фокусируют свое внимание на соответствующей задаче информации («теория контролируемого внимания рабочей памяти», например, Engle и др., 1999; Конвей и др., 2001; Кейн и Энгл, 2003; Энгл и Кейн, 2004). Например, при повторении услышанного слухового сообщения при одновременном игнорировании не относящегося к делу сообщения люди с низким объемом рабочей памяти в три раза чаще, чем люди с высоким объемом памяти, сообщают, что слышали свое имя в нерелевантном сообщении (Conway et al., 2001). Люди с высоким объемом рабочей памяти также быстрее, чем люди с более низким объемом памяти, идентифицируют целевую букву в незанятом месте, что подразумевает лучший контроль внимания у людей с высоким объемом памяти (Kane et al., 2001). Производительность у людей с высоким размахом по сравнению с людьми с низким размахом меньше зависит от присутствия неожиданного слухового отклонения, и их ERP также показывают меньший компонент N1, связанный с нечастыми слуховыми стимулами, что снова предполагает, что они лучше предотвращают обработку нерелевантных отвлекающих факторов. (Сорквист и др., 2012; Цучида и др., 2012). Многие косвенные доказательства связи между объемом рабочей памяти и уровнем вмешательства, создаваемого нерелевантными отвлекающими факторами, были получены в исследованиях когнитивного старения, которые, как правило, показывают, что пожилые участники непропорционально хуже выполняют задачи, требующие активного отказа от отвлекающей информации, по сравнению с более молодыми участниками. (Baddeley, 1996; De Fockert, 2005; De Fockert et al., 2009). Такие доказательства связи между объемом рабочей памяти и избирательным вниманием в основном косвенные, такие как снижение объема рабочей памяти у пожилых людей (по сравнению с взрослыми).более молодые) группы часто предполагается (например, Welford, 1958), а не измеряется в этих исследованиях.
(Сорквист и др., 2012; Цучида и др., 2012). Многие косвенные доказательства связи между объемом рабочей памяти и уровнем вмешательства, создаваемого нерелевантными отвлекающими факторами, были получены в исследованиях когнитивного старения, которые, как правило, показывают, что пожилые участники непропорционально хуже выполняют задачи, требующие активного отказа от отвлекающей информации, по сравнению с более молодыми участниками. (Baddeley, 1996; De Fockert, 2005; De Fockert et al., 2009). Такие доказательства связи между объемом рабочей памяти и избирательным вниманием в основном косвенные, такие как снижение объема рабочей памяти у пожилых людей (по сравнению с взрослыми).более молодые) группы часто предполагается (например, Welford, 1958), а не измеряется в этих исследованиях.
В ряде исследований связь между объемом рабочей памяти и обработкой отвлекающих факторов при избирательном внимании изучалась более непосредственно, путем измерения продолжительности рабочей памяти у молодых участников и сравнения производительности избирательного внимания между группами с низкой и высокой продолжительностью. Измерение объема рабочей памяти человека обычно получают с помощью стандартной задачи на диапазон, такой как задача Operation Span (Ospan) (Unsworth et al., 2005). В Ospan участники выполняют числовые вычисления, добавляя к списку слов, которые они хранят в памяти, а объем рабочей памяти представляет собой сумму всех правильно воспроизведенных списков слов. В стандартной задаче Струпа, требующей названия цвета чернил напечатанных строк букв (Stroop, 1935), интерференционные эффекты, возникающие, когда строка букв читает название другого цвета, постоянно выше у людей с низким объемом рабочей памяти по сравнению с людьми с высоким объемом рабочей памяти (Kane). и Энгл, 2003). Более сильные интерференционные эффекты также проявляются во фланкерных задачах типа Эриксена на низких (по сравнению сhigh) охватывают особей (Ahmed, De Fockert, 2012b; Shipstead et al., 2012). В совокупности эти данные свидетельствуют о том, что низкий объем рабочей памяти оказывает такое же влияние на отвлекаемость, как и высокая нагрузка на рабочую память.
Измерение объема рабочей памяти человека обычно получают с помощью стандартной задачи на диапазон, такой как задача Operation Span (Ospan) (Unsworth et al., 2005). В Ospan участники выполняют числовые вычисления, добавляя к списку слов, которые они хранят в памяти, а объем рабочей памяти представляет собой сумму всех правильно воспроизведенных списков слов. В стандартной задаче Струпа, требующей названия цвета чернил напечатанных строк букв (Stroop, 1935), интерференционные эффекты, возникающие, когда строка букв читает название другого цвета, постоянно выше у людей с низким объемом рабочей памяти по сравнению с людьми с высоким объемом рабочей памяти (Kane). и Энгл, 2003). Более сильные интерференционные эффекты также проявляются во фланкерных задачах типа Эриксена на низких (по сравнению сhigh) охватывают особей (Ahmed, De Fockert, 2012b; Shipstead et al., 2012). В совокупности эти данные свидетельствуют о том, что низкий объем рабочей памяти оказывает такое же влияние на отвлекаемость, как и высокая нагрузка на рабочую память.
Несколько исследований не смогли найти четких доказательств того, что низкий объем рабочей памяти всегда связан со сниженной избирательностью внимания. В недавнем исследовании только одна треть самоотчетов о ежедневных нарушениях внимания показала значительную корреляцию с объемом рабочей памяти (Unsworth et al., 2012), хотя сообщаемые нарушения внимания в основном связаны с рассеянностью и блужданием ума, а не с мерами избирательного внимания. Другое недавнее исследование показало, что более высокая отвлекаемость у людей с низким (по сравнению с высоким) объемом памяти может быть обращена вспять, если одновременно манипулировать нагрузкой на рабочую память (Ahmed and De Fockert, 2012b). В то время как эффекты фланговой интерференции были сильнее у людей с низким (по сравнению с высоким) объемом памяти, когда нагрузка на одновременную задачу на рабочую память была низкой, интерференционные эффекты показали противоположную картину при высокой нагрузке на рабочую память, так что люди с низким объемом памяти меньше отвлекались, чем люди с высоким объемом памяти. охватывать.Мы вернемся к этому открытию в следующем разделе, когда будем обсуждать возможные механизмы, с помощью которых рабочая память может влиять на избирательное внимание.
охватывать.Мы вернемся к этому открытию в следующем разделе, когда будем обсуждать возможные механизмы, с помощью которых рабочая память может влиять на избирательное внимание.
Таким образом, представление о том, что рабочая память играет роль в избирательном внимании, хорошо подтверждается исследованиями индивидуальных различий в рабочей памяти. Возможно, в большей степени, чем манипуляции с доступностью рабочей памяти внутри участников, которые дали некоторые противоречивые результаты, индивидуальные различия в объеме рабочей памяти довольно последовательно связаны с различиями в избирательном внимании, так что низкий объем рабочей памяти связан со снижением производительности. на заданиях на избирательное внимание, включая задания, связанные с игнорированием потенциально отвлекающей информации.Мета-анализ величины эффекта, о котором сообщалось в исследованиях по манипулированию измерением избирательного внимания в зависимости от объема рабочей памяти, показывает, что высокий объем рабочей памяти связан с уменьшением помех от дистракторов [среднее r = 0,286, t (11) = 7,92 , SEM = 0,0361, p < 0,001; см. табл. 2]. Этот вывод согласуется с теорией нагрузки избирательного внимания (Lavie et al., 2004), которая предсказывает, что любое снижение доступности рабочей памяти, будь то из-за высокой одновременной нагрузки на рабочую память, как обсуждалось в предыдущем разделе, или низкий объем рабочей памяти, как указано в текущем разделе, поставит под угрозу способность эффективно контролировать обработку воспринимаемых отвлекающих факторов.
табл. 2]. Этот вывод согласуется с теорией нагрузки избирательного внимания (Lavie et al., 2004), которая предсказывает, что любое снижение доступности рабочей памяти, будь то из-за высокой одновременной нагрузки на рабочую память, как обсуждалось в предыдущем разделе, или низкий объем рабочей памяти, как указано в текущем разделе, поставит под угрозу способность эффективно контролировать обработку воспринимаемых отвлекающих факторов.
Таблица 2. Величина эффекта ( r ) влияния объема рабочей памяти на обработку дистрактора (влияние времени реакции, если не указано иное; высокий и низкий балл по показателю объема рабочей памяти, если не указано иное) .
Возможные механизмы, лежащие в основе связи между рабочей памятью и избирательным вниманием
Хотя имеющиеся данные ясно указывают на степень функционального перекрытия между рабочей памятью и избирательным вниманием, до недавнего времени точная природа взаимодействия между рабочей памятью и избирательным вниманием оставалась относительно неясной. Связь между рабочей памятью и избирательным вниманием может просто заключаться в использовании одного и того же ограниченного пула ресурсов, необходимого как для активного хранения информации в рабочей памяти, так и для активного подавления отвлекающих факторов при избирательном внимании. С другой стороны, рабочая память может играть более специфическую роль в избирательном внимании, например, сохраняя четкие приоритеты для обработки информации, связанной с целью, при избирательном внимании (Lavie et al., 2004). В этом разделе будут обсуждаться два вопроса.Рабочая память — это многокомпонентная система, и первый вопрос касается того, участвуют ли какие-либо конкретные компоненты рабочей памяти в избирательном внимании, и если да, то какими могут быть эти компоненты. Во-вторых, хотя влияние рабочей памяти на избирательное внимание хорошо задокументировано с точки зрения простой степени обработки отвлекающей информации, остается неясным, какой функциональный механизм избирательного внимания лежит в основе этого эффекта.
Связь между рабочей памятью и избирательным вниманием может просто заключаться в использовании одного и того же ограниченного пула ресурсов, необходимого как для активного хранения информации в рабочей памяти, так и для активного подавления отвлекающих факторов при избирательном внимании. С другой стороны, рабочая память может играть более специфическую роль в избирательном внимании, например, сохраняя четкие приоритеты для обработки информации, связанной с целью, при избирательном внимании (Lavie et al., 2004). В этом разделе будут обсуждаться два вопроса.Рабочая память — это многокомпонентная система, и первый вопрос касается того, участвуют ли какие-либо конкретные компоненты рабочей памяти в избирательном внимании, и если да, то какими могут быть эти компоненты. Во-вторых, хотя влияние рабочей памяти на избирательное внимание хорошо задокументировано с точки зрения простой степени обработки отвлекающей информации, остается неясным, какой функциональный механизм избирательного внимания лежит в основе этого эффекта.
Какие компоненты рабочей памяти могут участвовать в избирательном внимании? Во многих исследованиях роли рабочей памяти в избирательном внимании использовались вербальные задания на рабочую память для манипулирования нагрузкой (т.г., Лави и др., 2004). Тот факт, что нагрузка на вербальную рабочую память приводит к усилению обработки отвлекающих факторов при избирательном внимании, может указывать на фонологическую петлю системы рабочей памяти как на ключевую подсистему, особенно потому, что задача невербальной рабочей памяти, такая как рабочая память на пространственные местоположения, делает это. не всегда имеют одинаковый эффект увеличения отвлекаемости при избирательном внимании (Kim et al., 2005; но см. Stins et al., 2004, доказательства того, что высокая нагрузка на пространственную рабочую память приводит к большему вмешательству Струпа).Два вывода, однако, опровергают этот вывод. Во-первых, эффект нагрузки рабочей памяти на отвлекаемость сохраняется даже тогда, когда фонологическая петля загружена явным повторением как при высокой, так и при низкой нагрузке (Lavie et al. , 2004, Experiment 2). Во-вторых, манипулирование нагрузкой когнитивного контроля, отличной от вербальной рабочей памяти, также приводит к повышенной отвлекаемости (Lavie et al., 2004; De Fockert et al., 2010). Отвлекающие эффекты сильнее, когда есть цена одновременного выполнения (Navon and Gopher, 1979), например, когда задача включает переключение между рабочей памятью и задачей избирательного внимания, по сравнению с тем, когда задача избирательного внимания выполняется сама по себе (Lavie и другие., 2004, Эксперименты 4 и 5). Точно так же время от времени необходимость давать пространственно неконгруэнтную реакцию, удерживая при этом пространственно преобладающую конгруэнтную реакцию, которая требуется в большинстве испытаний (манипуляция, которая не нагружает рабочую память), также приводит к повышенной отвлекаемости при последующем фланговом задании (Де Фоккерт). и др., 2010).
, 2004, Experiment 2). Во-вторых, манипулирование нагрузкой когнитивного контроля, отличной от вербальной рабочей памяти, также приводит к повышенной отвлекаемости (Lavie et al., 2004; De Fockert et al., 2010). Отвлекающие эффекты сильнее, когда есть цена одновременного выполнения (Navon and Gopher, 1979), например, когда задача включает переключение между рабочей памятью и задачей избирательного внимания, по сравнению с тем, когда задача избирательного внимания выполняется сама по себе (Lavie и другие., 2004, Эксперименты 4 и 5). Точно так же время от времени необходимость давать пространственно неконгруэнтную реакцию, удерживая при этом пространственно преобладающую конгруэнтную реакцию, которая требуется в большинстве испытаний (манипуляция, которая не нагружает рабочую память), также приводит к повышенной отвлекаемости при последующем фланговом задании (Де Фоккерт). и др., 2010).
Выводы о том, что манипуляции, которые задействуют функции когнитивного контроля, отличные от рабочей памяти, такие как выполнение двойной задачи и торможение реакции, также связаны с усилением обработки отвлекающих факторов при избирательном внимании, предполагают, что эффект рабочей памяти при избирательном внимании более вероятен. быть общим для предметной области, чем специфичной для предметной области (De Fockert and Bremner, 2011).Обобщенный характер роли рабочей памяти в избирательном внимании дополнительно демонстрируется тем фактом, что нагрузка на вербальную рабочую память влияет на обработку нерелевантной информации в задачах, которые вряд ли полагаются на активацию вербальных кодов, таких как визуальный поиск в памяти. исследования захвата внимания (Lavie and De Fockert, 2005), а также суждения о размере при иллюзии Эббингауза (De Fockert and Wu, 2009) и слепоте по невнимательности (De Fockert and Bremner, 2011). Очевидным кандидатом на роль общего компонента рабочей памяти, участвующего в избирательном внимании, является центральный исполнительный орган.Действительно, предполагалось, что одной из первоначальных функций центрального исполнительного органа будет избирательное внимание к важной для задачи информации перед лицом потенциального отвлечения внимания другими источниками (Baddeley, 1996). Центральный исполнительный орган, однако, не играет прямой роли в хранении данных, и хранилищем часто манипулируют для увеличения нагрузки на рабочую память. Наряду с открытием того, что манипуляции, нагружающие функции когнитивного контроля, отличные от рабочей памяти, включая выполнение двойных задач и подавление доминантных реакций, также усиливают обработку отвлекающих факторов при избирательном внимании, это оставляет открытой возможность того, что рабочая память оказывает косвенное влияние на избирательное внимание, возможно, потому, что оно разделяет ограниченные ресурсы с механизмами когнитивного контроля, участвующими в активном отклонении отвлекающих факторов.Необходима дополнительная работа для дальнейшего изучения этой возможности.
быть общим для предметной области, чем специфичной для предметной области (De Fockert and Bremner, 2011).Обобщенный характер роли рабочей памяти в избирательном внимании дополнительно демонстрируется тем фактом, что нагрузка на вербальную рабочую память влияет на обработку нерелевантной информации в задачах, которые вряд ли полагаются на активацию вербальных кодов, таких как визуальный поиск в памяти. исследования захвата внимания (Lavie and De Fockert, 2005), а также суждения о размере при иллюзии Эббингауза (De Fockert and Wu, 2009) и слепоте по невнимательности (De Fockert and Bremner, 2011). Очевидным кандидатом на роль общего компонента рабочей памяти, участвующего в избирательном внимании, является центральный исполнительный орган.Действительно, предполагалось, что одной из первоначальных функций центрального исполнительного органа будет избирательное внимание к важной для задачи информации перед лицом потенциального отвлечения внимания другими источниками (Baddeley, 1996). Центральный исполнительный орган, однако, не играет прямой роли в хранении данных, и хранилищем часто манипулируют для увеличения нагрузки на рабочую память. Наряду с открытием того, что манипуляции, нагружающие функции когнитивного контроля, отличные от рабочей памяти, включая выполнение двойных задач и подавление доминантных реакций, также усиливают обработку отвлекающих факторов при избирательном внимании, это оставляет открытой возможность того, что рабочая память оказывает косвенное влияние на избирательное внимание, возможно, потому, что оно разделяет ограниченные ресурсы с механизмами когнитивного контроля, участвующими в активном отклонении отвлекающих факторов.Необходима дополнительная работа для дальнейшего изучения этой возможности.
Какие функциональные механизмы избирательного внимания могут объяснить наблюдаемые изменения отвлекаемости в зависимости от наличия рабочей памяти? Было предложено по крайней мере две возможности, основанные либо на временных, либо на пространственных аспектах избирательного внимания. Высокая нагрузка на рабочую память приводит к задержке выделения внимания на целевые представления при обработке зрительной информации (Scalf et al. , 2011): нейронный ответ в затылочной коре, связанный со зрительной мишенью, достигает максимума позже при нагрузке на одновременную задачу рабочей памяти. был высоким (по сравнению снизкая), что свидетельствует о том, что зрительное внимание медленнее направляется к соответствующему целевому представлению, когда рабочая память задействована иным образом. Другие результаты подтверждают мнение о том, что доступность рабочей памяти влияет на синхронизацию процессов внимания (например, Heitz and Engle, 2007; Poole and Kane, 2009). Во время обработки фланкерных проб люди с низким и высоким объемом рабочей памяти изначально одинаково отвлекаются на нерелевантную информацию, поскольку они показывают одинаковую точность для ответов быстрее, чем ~ 400 мс.Однако производительность улучшается быстрее у людей с высоким (по сравнению с низким) размахом, поскольку ответы становятся медленнее, что может свидетельствовать о том, что люди с высоким размахом быстрее концентрируют внимание на соответствующей информации (Heitz and Engle, 2007).
, 2011): нейронный ответ в затылочной коре, связанный со зрительной мишенью, достигает максимума позже при нагрузке на одновременную задачу рабочей памяти. был высоким (по сравнению снизкая), что свидетельствует о том, что зрительное внимание медленнее направляется к соответствующему целевому представлению, когда рабочая память задействована иным образом. Другие результаты подтверждают мнение о том, что доступность рабочей памяти влияет на синхронизацию процессов внимания (например, Heitz and Engle, 2007; Poole and Kane, 2009). Во время обработки фланкерных проб люди с низким и высоким объемом рабочей памяти изначально одинаково отвлекаются на нерелевантную информацию, поскольку они показывают одинаковую точность для ответов быстрее, чем ~ 400 мс.Однако производительность улучшается быстрее у людей с высоким (по сравнению с низким) размахом, поскольку ответы становятся медленнее, что может свидетельствовать о том, что люди с высоким размахом быстрее концентрируют внимание на соответствующей информации (Heitz and Engle, 2007).
Рабочая память также может влиять на то, как избирательное внимание работает в пространственной области. Доказательства того, что люди с высоким уровнем рабочей памяти быстрее, чем люди с низким объемом рабочей памяти, ограничивают пространственное внимание значимыми для задачи местами, тем самым исключая места отвлекающих факторов из обработки (Heitz and Engle, 2007; Poole and Kane, 2009), предполагают, что как временные, так и пространственные аспекты избирательного внимания зависят от рабочей памяти.Также было показано, что высокая емкость рабочей памяти позволяет лучше ограничивать пространственное внимание в течение длительного периода времени (Poole and Kane, 2009). Еще одно доказательство того, что нагрузка на рабочую память влияет на распределение пространственного внимания, получено в исследовании с использованием фланговой задачи, в которой интерференция дистрактора измерялась при различных пространственных расстояниях между целью и дистрактором при низкой или высокой нагрузке на рабочую память (Капарос и Линнелл). , 2010). Высокая нагрузка привела к диспергированию характерного пространственного профиля внимания с точки зрения зон фасилитации и подавления (Müller et al., 2005). Аналогичные результаты были получены при выполнении фланкерной задачи при одновременном манипулировании факторами нагрузки на рабочую память и объемом рабочей памяти (Ahmed and De Fockert, 2012b). Пространственный профиль внимания был наиболее ограничен у лиц с высоким объемом рабочей памяти, в то время как нагрузка на задачу рабочей памяти была низкой. Подобные и более рассеянные профили, чем в группе с большим пролетом при низкой нагрузке, наблюдались при высокой нагрузке в группе с большим пролетом и при низкой нагрузке в группе с низким пролетом.Когда рабочая память была высокой в группе с низким объемом памяти, пространственное внимание становилось еще менее ограниченным. Все эти результаты свидетельствуют о том, что рабочая память необходима для поддержания узкого фокуса внимания, соответствующего задаче, и что недоступность рабочей памяти приводит к расширению фокуса внимания.
, 2010). Высокая нагрузка привела к диспергированию характерного пространственного профиля внимания с точки зрения зон фасилитации и подавления (Müller et al., 2005). Аналогичные результаты были получены при выполнении фланкерной задачи при одновременном манипулировании факторами нагрузки на рабочую память и объемом рабочей памяти (Ahmed and De Fockert, 2012b). Пространственный профиль внимания был наиболее ограничен у лиц с высоким объемом рабочей памяти, в то время как нагрузка на задачу рабочей памяти была низкой. Подобные и более рассеянные профили, чем в группе с большим пролетом при низкой нагрузке, наблюдались при высокой нагрузке в группе с большим пролетом и при низкой нагрузке в группе с низким пролетом.Когда рабочая память была высокой в группе с низким объемом памяти, пространственное внимание становилось еще менее ограниченным. Все эти результаты свидетельствуют о том, что рабочая память необходима для поддержания узкого фокуса внимания, соответствующего задаче, и что недоступность рабочей памяти приводит к расширению фокуса внимания. Действительно, влияние нагрузки рабочей памяти на отвлекаемость отсутствует, когда фокус внимания экспериментально манипулируют, чтобы он оставался либо узким, либо широким (Chen and Chan, 2007).
Действительно, влияние нагрузки рабочей памяти на отвлекаемость отсутствует, когда фокус внимания экспериментально манипулируют, чтобы он оставался либо узким, либо широким (Chen and Chan, 2007).
Совместное влияние нагрузки на рабочую память и объема памяти на пространственный профиль внимания может объяснить кажущиеся противоречивыми выводы о том, что нагрузка на рабочую память может иметь противоположные эффекты в зависимости от объема рабочей памяти человека, увеличивая отвлекаемость у людей с высоким объемом рабочей памяти, но уменьшая отвлекаемость у людей с низким размахом (Ahmed and De Fockert, 2012b).Пространственный профиль внимания состоит из чередующихся зон фасилитации и подавления (Müller et al., 2005), а закономерность эффектов дистрактора в зависимости как от нагрузки, так и от емкости рабочей памяти точно объясняется в терминах области профиля внимания (Müller et al., 2005). фасилитация, подавление), с которым совпадает дистрактор, давая предполагаемые изменения пространственной дисперсии профиля внимания в зависимости от нагрузки и объема рабочей памяти. Подобные изменения в пространственном профиле внимания также могут объяснить тот факт, что интерференционные эффекты на глобальном уровне фигуры Навона усиливаются, но интерференционные эффекты на локальном уровне уменьшаются из-за высокой нагрузки на рабочую память (Ahmed and De Fockert, 2012a).Более рассредоточенный профиль внимания при высокой нагрузке повысит вероятность того, что будет задействован глобальный уровень фигуры Навона, что приведет к большему вмешательству, когда внимание должно быть направлено на локальный уровень, и меньшему вмешательству, когда оно должно быть направлено на глобальный уровень. уровень.
Подобные изменения в пространственном профиле внимания также могут объяснить тот факт, что интерференционные эффекты на глобальном уровне фигуры Навона усиливаются, но интерференционные эффекты на локальном уровне уменьшаются из-за высокой нагрузки на рабочую память (Ahmed and De Fockert, 2012a).Более рассредоточенный профиль внимания при высокой нагрузке повысит вероятность того, что будет задействован глобальный уровень фигуры Навона, что приведет к большему вмешательству, когда внимание должно быть направлено на локальный уровень, и меньшему вмешательству, когда оно должно быть направлено на глобальный уровень. уровень.
Таким образом, влияние рабочей памяти на избирательное внимание в значительной степени носит общий характер, и рабочая память и избирательное внимание взаимодействуют даже тогда, когда задачи мало перекрываются с точки зрения содержания стимула (но см. Kim et al., 2005; Park et al., 2007, за результаты, свидетельствующие о доменно-специфических эффектах рабочей памяти на выбор). Более того, существует по крайней мере два возможных способа влияния рабочей памяти на избирательное внимание. Во-первых, временная динамика выбора внимания может измениться, когда рабочая память недоступна для выбора. При высокой нагрузке на рабочую память активация представления визуальной цели задерживается, что дает больше возможностей для влияния отвлекающих факторов на поведение. Точно так же низкий объем рабочей памяти связан с более медленным ограничением внимания входными данными, относящимися к задаче, что опять же оставляет более длительное временное окно, в котором могут обрабатываться отвлекающие факторы.Во-вторых, пространственное распределение внимания зависит от наличия рабочей памяти. Пространственное внимание имеет более ограниченный фокус, когда рабочая память доступна для внимания (при низкой нагрузке на рабочую память или у людей с большим объемом рабочей памяти), по сравнению с тем, когда рабочая память менее доступна (при высокой нагрузке на рабочую память или у людей с малый объем рабочей памяти).
Более того, существует по крайней мере два возможных способа влияния рабочей памяти на избирательное внимание. Во-первых, временная динамика выбора внимания может измениться, когда рабочая память недоступна для выбора. При высокой нагрузке на рабочую память активация представления визуальной цели задерживается, что дает больше возможностей для влияния отвлекающих факторов на поведение. Точно так же низкий объем рабочей памяти связан с более медленным ограничением внимания входными данными, относящимися к задаче, что опять же оставляет более длительное временное окно, в котором могут обрабатываться отвлекающие факторы.Во-вторых, пространственное распределение внимания зависит от наличия рабочей памяти. Пространственное внимание имеет более ограниченный фокус, когда рабочая память доступна для внимания (при низкой нагрузке на рабочую память или у людей с большим объемом рабочей памяти), по сравнению с тем, когда рабочая память менее доступна (при высокой нагрузке на рабочую память или у людей с малый объем рабочей памяти).
Обсуждение
Идея о том, что рабочая память и избирательное внимание являются тесно связанными системами, стала общепринятой в последнее десятилетие до такой степени, что их иногда рассматривают как два проявления одной и той же основной системы (Awh and Jonides, 2001; Awh et al., 2006; Чун, 2011). Как рабочая память, так и избирательное внимание включают в себя определение приоритетов определенной информации при наличии конкурирующих входных данных и включают сохранение информации во времени (рабочая память) и в пространстве (избирательное внимание). Кроме того, рабочая память и избирательное внимание имеют общие нейронные системы (например, Gazzaley et al., 2007; Mayer et al., 2007; Gazzaley and Nobre, 2012), и результаты людей в задачах на рабочую память и избирательное внимание демонстрируют устойчивую положительную корреляцию ( е.г., Кейн и Энгл, 2003). Рассмотренная здесь работа с достаточной последовательностью обнаружила, что недоступность рабочей памяти для избирательного внимания либо потому, что рабочая память задействована в дополнительной задаче с высокой нагрузкой, либо из-за низкой емкости рабочей памяти, связана с большей отвлекаемостью в диапазоне задания на избирательное внимание. Этот вывод подтверждается мета-анализом размеров эффекта в исследованиях манипулирования нагрузкой на рабочую память во время избирательного внимания, который показал, что высокая нагрузка на рабочую память имеет тенденцию приводить к увеличению обработки отвлекающих факторов.
Этот вывод подтверждается мета-анализом размеров эффекта в исследованиях манипулирования нагрузкой на рабочую память во время избирательного внимания, который показал, что высокая нагрузка на рабочую память имеет тенденцию приводить к увеличению обработки отвлекающих факторов.
Альтернативное объяснение наблюдаемого влияния рабочей памяти на избирательное внимание заключается в том, что увеличение отвлекаемости при высокой нагрузке на рабочую память происходит просто из-за увеличения общей сложности задачи при высокой нагрузке на рабочую память. Две линии доказательств выступают против этого аргумента. Во-первых, когда увеличивается общая задержка ответа и/или частота ошибок при высокой нагрузке на рабочую память, что предполагает общее увеличение сложности задачи, увеличение эффекта отвлекающих помех имеет тенденцию быть непропорциональным увеличению общей стоимости производительности. е.г., Лави и др., 2004). Во-вторых, производительность, которая выигрывает от большей обработки отвлекающих факторов, должна быть выше при высокой нагрузке на рабочую память. Есть несколько примеров того, что это действительно так. Негативный прайминг означает обнаружение нарушения обработки недавно проигнорированной информации (Типпер, 1985). Следовательно, если отвлекающие факторы нельзя эффективно игнорировать при высокой нагрузке на рабочую память, то негативные эффекты прайминга должны быть уменьшены при высокой нагрузке. Это действительно так, и ухудшение производительности обычно наблюдается, когда ранее игнорируемый отвлекающий фактор повторяется по мере того, как цель устраняется из-за высокой нагрузки на рабочую память (De Fockert et al., 2010). Другим примером улучшения производительности за счет высокой нагрузки на рабочую память является освобождение от слепоты по невнимательности, обнаруженное при измерении показателей обнаружения и идентификации неожиданного, не относящегося к задаче визуального стимула при различных уровнях нагрузки на рабочую память (De Fockert and Bremner, 2011). .
Есть несколько примеров того, что это действительно так. Негативный прайминг означает обнаружение нарушения обработки недавно проигнорированной информации (Типпер, 1985). Следовательно, если отвлекающие факторы нельзя эффективно игнорировать при высокой нагрузке на рабочую память, то негативные эффекты прайминга должны быть уменьшены при высокой нагрузке. Это действительно так, и ухудшение производительности обычно наблюдается, когда ранее игнорируемый отвлекающий фактор повторяется по мере того, как цель устраняется из-за высокой нагрузки на рабочую память (De Fockert et al., 2010). Другим примером улучшения производительности за счет высокой нагрузки на рабочую память является освобождение от слепоты по невнимательности, обнаруженное при измерении показателей обнаружения и идентификации неожиданного, не относящегося к задаче визуального стимула при различных уровнях нагрузки на рабочую память (De Fockert and Bremner, 2011). .
Теория нагрузки предсказывает противоположное влияние на отвлекаемость в зависимости от увеличения нагрузки на перцепцию и рабочей памяти (Lavie et al. , 2004). Однако вопрос о том, можно ли четко различать конструкты нагрузки на перцепцию и нагрузки на рабочую память, был поставлен под сомнение, и было высказано предположение, что предсказание повышенной отвлекаемости при высокой нагрузке на рабочую память справедливо только в том случае, если нагрузка на перцепцию и рабочую память независимы и зависят от отдельных факторов. ресурсы (Цал и Бенони, 2010).Если этого не происходит, то любое дополнительное увеличение нагрузки на ресурсы внимания (перцептивные или когнитивные) должно быть связано со снижением обработки отвлекающих факторов. Данные исследований нагрузки на рабочую память и объема рабочей памяти явно не подтверждают эту интерпретацию теории нагрузки, предполагая, что когнитивную и перцептивную нагрузку следует рассматривать как отдельные конструкции. Действительно, в то время как пространственная рабочая память и пространственное избирательное внимание могут совместно использовать определенные ресурсы (Awh and Jonides, 2001), существует меньше свидетельств какой-либо прямой конкуренции за ресурсы между обсуждаемыми здесь типами рабочей памяти (например,г.
, 2004). Однако вопрос о том, можно ли четко различать конструкты нагрузки на перцепцию и нагрузки на рабочую память, был поставлен под сомнение, и было высказано предположение, что предсказание повышенной отвлекаемости при высокой нагрузке на рабочую память справедливо только в том случае, если нагрузка на перцепцию и рабочую память независимы и зависят от отдельных факторов. ресурсы (Цал и Бенони, 2010).Если этого не происходит, то любое дополнительное увеличение нагрузки на ресурсы внимания (перцептивные или когнитивные) должно быть связано со снижением обработки отвлекающих факторов. Данные исследований нагрузки на рабочую память и объема рабочей памяти явно не подтверждают эту интерпретацию теории нагрузки, предполагая, что когнитивную и перцептивную нагрузку следует рассматривать как отдельные конструкции. Действительно, в то время как пространственная рабочая память и пространственное избирательное внимание могут совместно использовать определенные ресурсы (Awh and Jonides, 2001), существует меньше свидетельств какой-либо прямой конкуренции за ресурсы между обсуждаемыми здесь типами рабочей памяти (например,г. , рабочая память на цифры) и зрительный выбор. Например, неперекрывающиеся области коры участвуют в задачах рабочей памяти для порядка цифр и зрительного выбора (например, De Fockert et al., 2001). Кроме того, неоднократно показано, что нагрузка на перцепцию и нагрузка на рабочую память оказывают противоположное влияние на отбор в исследованиях нагрузки на перцепцию или на рабочую память, а также, когда в одном и том же эксперименте манипулируют обоими типами нагрузки, обработка дистракторами снижается, когда нагрузка на перцепцию снижается. высоким, но увеличивается при высокой нагрузке на рабочую память (Lavie et al., 2004, Эксперимент 3). В другой работе была обнаружена аналогичная диссоциация между эффектами двух типов нагрузки на обработку дистракторов (Yi et al., 2004).
, рабочая память на цифры) и зрительный выбор. Например, неперекрывающиеся области коры участвуют в задачах рабочей памяти для порядка цифр и зрительного выбора (например, De Fockert et al., 2001). Кроме того, неоднократно показано, что нагрузка на перцепцию и нагрузка на рабочую память оказывают противоположное влияние на отбор в исследованиях нагрузки на перцепцию или на рабочую память, а также, когда в одном и том же эксперименте манипулируют обоими типами нагрузки, обработка дистракторами снижается, когда нагрузка на перцепцию снижается. высоким, но увеличивается при высокой нагрузке на рабочую память (Lavie et al., 2004, Эксперимент 3). В другой работе была обнаружена аналогичная диссоциация между эффектами двух типов нагрузки на обработку дистракторов (Yi et al., 2004).
В настоящее время начинают решать ряд ключевых вопросов о роли рабочей памяти в избирательном внимании. Во-первых, кажется, что рабочая память влияет на выбор на ранней стадии обработки, часто влияя на восприятие отвлекающей информации (например, De Fockert and Bremner, 2011). Во-вторых, влияние рабочей памяти на избирательное внимание носит общий характер, так как загрузка рабочей памяти в одном домене (т.g., сохранение визуально представленных цифр) может привести к усилению обработки отвлекающих факторов в другой области (например, обработка одиночного цвета или отвлекающего круга; например, Lavie and De Fockert, 2005; De Fockert and Wu, 2009) или даже в другой модальности (например, прослушивание или прикосновение; Dalton et al., 2009a,b). Более того, другие манипуляции когнитивного контроля (например, выполнение двойной задачи и подавление реакции) влияют на обработку дистракторов так же, как загрузка рабочей памяти. В-третьих, нагрузка на рабочую память может влиять как на временное, так и на пространственное развертывание внимания (Caparos and Linnell, 2010; Scalf et al., 2011; Ахмед и Де Фокерт, 2012b).
Во-вторых, влияние рабочей памяти на избирательное внимание носит общий характер, так как загрузка рабочей памяти в одном домене (т.g., сохранение визуально представленных цифр) может привести к усилению обработки отвлекающих факторов в другой области (например, обработка одиночного цвета или отвлекающего круга; например, Lavie and De Fockert, 2005; De Fockert and Wu, 2009) или даже в другой модальности (например, прослушивание или прикосновение; Dalton et al., 2009a,b). Более того, другие манипуляции когнитивного контроля (например, выполнение двойной задачи и подавление реакции) влияют на обработку дистракторов так же, как загрузка рабочей памяти. В-третьих, нагрузка на рабочую память может влиять как на временное, так и на пространственное развертывание внимания (Caparos and Linnell, 2010; Scalf et al., 2011; Ахмед и Де Фокерт, 2012b).
В заключение, в этой статье представлен обзор доказательств формы выбора внимания, которая отличается от типа выборочной обработки, основанной на перцептивных аспектах ввода, предложенных моделями перцептивной нагрузки и разбавления. В то время как отбор часто может происходить пассивно из-за характеристик входных данных (т. е. в условиях высокой перцептивной нагрузки или сильного разбавления), отвлекающая информация часто подвергается некоторой обработке и требует активного отбора, например, когда отвлекающие факторы достаточно заметны или непредвиденный.В таких случаях потенциального отвлечения рабочая память играет роль в минимизации помех, создаваемых отвлекающими факторами. Это всего лишь один из способов взаимодействия рабочей памяти и избирательного внимания, а другие связи включают роль избирательного внимания в определении того, какая информация закодирована в рабочей памяти (например, Oberauer, 2003), и обнаружение того, что содержимое рабочей памяти может влиять на то, что выбирается при визуальной обработке (Soto et al., 2008). Эти множественные взаимодействия подчеркивают тесную связь между рабочей памятью и избирательным вниманием.
В то время как отбор часто может происходить пассивно из-за характеристик входных данных (т. е. в условиях высокой перцептивной нагрузки или сильного разбавления), отвлекающая информация часто подвергается некоторой обработке и требует активного отбора, например, когда отвлекающие факторы достаточно заметны или непредвиденный.В таких случаях потенциального отвлечения рабочая память играет роль в минимизации помех, создаваемых отвлекающими факторами. Это всего лишь один из способов взаимодействия рабочей памяти и избирательного внимания, а другие связи включают роль избирательного внимания в определении того, какая информация закодирована в рабочей памяти (например, Oberauer, 2003), и обнаружение того, что содержимое рабочей памяти может влиять на то, что выбирается при визуальной обработке (Soto et al., 2008). Эти множественные взаимодействия подчеркивают тесную связь между рабочей памятью и избирательным вниманием.
Заявление о конфликте интересов
Автор заявляет, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Ссылки
Ахмед Л. и Де Фоккерт Дж. В. (2012a). Нагрузка на рабочую память может как улучшать, так и ухудшать избирательное внимание: свидетельство парадигмы Навона. Аттен. Восприятие. Психофиз . 74, 1397–1405.
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Ахмед, Л.и Де Фокерт, Дж. В. (2012b). Сосредоточение внимания: влияние объема рабочей памяти и нагрузки на избирательное внимание. PLoS ONE 7:e43101. doi: 10.1371/journal.pone.0043101
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Баддели, А. (1996). Изучение центральной исполнительной власти. QJ Exp. Психол . 49А, 5–28.
Бенони, Х., и Цал, Ю. (2012). Контроль разбавления при манипулировании нагрузкой: перцептивные и сенсорные ограничения — это всего лишь два аспекта сложности задачи. Психон. Бык. Версия . 19, 631–638.
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Бут, В. Р., Брокмоул, Дж. Р., и Саймонс, Д. Дж. (2005). Привлечение внимания модулируется в ситуациях с двумя задачами. Психон. Бык. Версия . 12, 662–668.
Р., Брокмоул, Дж. Р., и Саймонс, Д. Дж. (2005). Привлечение внимания модулируется в ситуациях с двумя задачами. Психон. Бык. Версия . 12, 662–668.
Опубликовано Резюме | Опубликован полный текст
Капарос, С., Ахмед, Л., Бремнер, А. Дж., Де Фокерт, Дж. В., Линнелл, К. Дж., и Давидофф, Дж. (2012). Воздействие городской среды изменяет местный уклон отдаленной культуры. Познание 122, 80–85.
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Капарос, С., Линнелл, К. Дж., Бремнер, А. Дж., Де Фокерт, Дж. В., и Давидофф, Дж. (2013). Рассказывают ли нам что-нибудь о локальном и глобальном избирательном внимании локальные и глобальные искажения восприятия? Психолог. Наука . 24, 206–212.
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Кармель, Д., Фэрни, Дж., и Лави, Н. (2012). Взвесьте и посмотрите: загрузка рабочей памяти улучшает случайную идентификацию нерелевантных лиц. Фронт. Психология 3:286. doi: 10.3389/fpsyg.2012.00286
Фронт. Психология 3:286. doi: 10.3389/fpsyg.2012.00286
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Чен, З., и Чан, К.С. (2007). Помехи дистрактора остаются постоянными, несмотря на изменение нагрузки на рабочую память. Психон. Бык. Версия . 14, 306–312.
Опубликовано Резюме | Опубликован полный текст
Конвей, А.Р.А., Коуэн, Н., и Бантинг, М.Ф. (2001). Новый взгляд на феномен коктейльной вечеринки: важность объема рабочей памяти. Психон. Бык. Версия . 8, 331–335.
Опубликовано Резюме | Опубликован полный текст
Де Фокерт, Дж., Рамчурн, А., Ван Вельзен, Дж., Бергстрем, З., и Банс, Д. (2009). Поведенческие и ERP свидетельствуют об усилении вмешательства в пожилом возрасте. Мозг Res . 1282, 67–73.
Де Фокерт, Дж. В., и Бремнер, А. Дж. (2011). Высвобождение слепоты по невниманию при высокой нагрузке на рабочую память: выяснение взаимосвязи между рабочей памятью и избирательным вниманием. Познание 121, 400–408.
Познание 121, 400–408.
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Де Фокерт, Дж. В., и Ву, С. (2009). Высокая нагрузка на рабочую память приводит к усилению иллюзии Эббингауза. евро. Дж. Когн. Психол . 21, 961–970.
де Лиано, Б., Умильта, К., Стаблум, Ф., Тебальди, Ф., и Кантагалло, А. (2010). Вмешательство дистрактора внимания может быть уменьшено одновременной нагрузкой на рабочую память у здоровых участников и у пациентов с черепно-мозговой травмой. Мозговой код . 74, 298–305.
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Энгл Р.В. и Кейн М.Дж. (2004). «Управленческое внимание, объем рабочей памяти и двухфакторная теория когнитивного контроля», в The Psychology of Learning and Motivation , Vol. 44, изд. Б. Росс (Нью-Йорк, штат Нью-Йорк: Acodemic Press), 145–199.
Энгл, Р. В., Кейн, М. Дж., и Тухольски, С. В. (1999). «Индивидуальные различия в объеме рабочей памяти и то, что они говорят нам о контролируемом внимании, общем подвижном интеллекте и функциях префронтальной коры», в «Модели рабочей памяти: механизмы активного обслуживания и исполнительного контроля» , под редакцией А.Мияке и П. Шах (Лондон: Cambridge Press), 102–134.
(1999). «Индивидуальные различия в объеме рабочей памяти и то, что они говорят нам о контролируемом внимании, общем подвижном интеллекте и функциях префронтальной коры», в «Модели рабочей памяти: механизмы активного обслуживания и исполнительного контроля» , под редакцией А.Мияке и П. Шах (Лондон: Cambridge Press), 102–134.
Газзали, А., Риссман, Дж., Куни, Дж., Рутман, А., Зайберт, Т., Клапп, В., и др. (2007). Функциональные взаимодействия между префронтальной и зрительной ассоциативной корой способствуют нисходящей модуляции зрительной обработки. Церебр. Кортекс 17, 125–135.
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Хашер Л. и Закс Р. Т. (1988). Рабочая память, понимание и старение: обзор и новый взгляд. Психолог. Учиться. Мотив . 22, 122–149.
Йонген, Э.М.М., и Йонкман, Л.М. (2011). Влияние одновременной загрузки рабочей памяти на обработку дистракторов и контроль интерференции в задаче Струпа с именным лицом. Психофизиология 48, 31–43.
Психофизиология 48, 31–43.
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Кейн, М.Дж., и Энгл, Р.В. (2003). Объем рабочей памяти и контроль внимания: вклад игнорирования цели, конкуренции в ответ и поставленной задачи на вмешательство Струпа. Дж. Экспл. Психол. Ген . 132, 47–70.
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Лави, Н. (2010). Внимание, отвлечение и когнитивный контроль при нагрузке. Курс. Реж. Психол. Наука . 19, 143–148.
Лави, Н., и Де Фоккерт, Дж. В. (2005). Роль рабочей памяти в захвате внимания. Психон. Бык. Версия . 12, 669–674.
Опубликовано Резюме | Опубликован полный текст
Лави, Н., и Де Фоккерт, Дж.В. (2006). Фронтальный контроль внимания Захват при зрительном поиске. Виз. Код . 14, 863–876.
Лави, Н., и Цал, Ю. (1994). Перцептивная нагрузка как основная детерминанта локуса выбора в зрительном внимании. Восприятие. Психофиз . 56, 183–197.
Восприятие. Психофиз . 56, 183–197.
Опубликовано Резюме | Опубликован полный текст
Мак, А., и Рок, И. (1998). Слепота по невнимательности . Лондон: Массачусетский технологический институт Press.
Майер, Дж. С., Биттнер, Р. А., Николич, Д., Bledowski, C., Goebel, R., и Linden, D.E. (2007). Общие нейронные субстраты для зрительной рабочей памяти и внимания. Нейроизображение 36, 441–453.
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Моррис, Р.Г., Гик, М.Л., и Крейк, Ф.И.М. (1988). Ресурсы обработки и возрастные различия в рабочей памяти. Мем. Код . 16, 362–366.
Опубликовано Резюме | Опубликован полный текст
Навон, Д. (1977). Лес перед деревьями: приоритет глобальных признаков в зрительном восприятии. Познан. Психол . 9, 353–383.
Навон, Д., и Гофер, Д. (1979). Об экономике системы обработки человека. Психолог. Версия . 86, 214–253.
86, 214–253.
Olivers, C.N., Peters, J., Houtkamp, R., and Roelfsema, P.R. (2011). Разные состояния зрительной рабочей памяти: когда она направляет внимание, а когда нет. Тенденции Cogn. Наука . 15, 327–334.
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Парк, С., Ким, М.С. и Чун М.М. (2007). Параллельная нагрузка на рабочую память может способствовать избирательному вниманию: свидетельство специализированной нагрузки. Дж. Экспл. Психол. Гум. Восприятие. Выполните . 33, 1062–1075.
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Пекчиненда, А., и Хейл, М. (2007). Роль нагрузки рабочей памяти на избирательное внимание к аффективно валентной информации. евро. Дж. Когн. Психол . 19, 898–909.
Пул, Б. Дж. и Кейн, М. Дж.(2009). Объем рабочей памяти предсказывает исполнительный контроль зрительного поиска среди отвлекающих факторов: влияние устойчивого и избирательного внимания. QJ Exp. Психол . 62, 1430–1454.
QJ Exp. Психол . 62, 1430–1454.
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Пратт, Н., Уиллоуби, А., и Свик, Д. (2011). Влияние нагрузки рабочей памяти на зрительное избирательное внимание: поведенческие и электрофизиологические данные. Фронт. Гум. Нейроски . 5:57. doi: 10.3389/fnhum.2011.00057
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Робертс, Б., Харрис, М. Г., и Йейтс, Т. А. (2005). Роли размера и расстояния индуктора в иллюзии Эббингауза (круги Титченера). Восприятие 34, 847–856.
Опубликовано Резюме | Опубликован полный текст
Рок И., Линнетт К.М., Грант П. и Мак А. (1992). Восприятие без внимания: результаты нового метода. Познан. Психол . 24, 502–534.
Опубликовано Резюме | Опубликован полный текст
Сан-Мигель, И., Corral, M.J., and Escera, C. (2008). При нагрузке рабочая память снижает отвлечение: поведенческие и электрофизиологические данные из слухо-зрительной парадигмы отвлечения. Дж. Когн. Нейроски . 20, 1131–1145.
Дж. Когн. Нейроски . 20, 1131–1145.
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Сёрквист, П., Нёстль, А., и Халин, Н. (2012). Объем рабочей памяти модулирует скорость привыкания: свидетельство кросс-модальной парадигмы слухового отвлечения. Психон. Бык. Версия .19, 245–250.
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Стинс, Дж. Ф., Воссе, С., Бумсма, Д. И., и де Геус, Э. Дж. (2004). О роли рабочей памяти в ответной интерференции. Восприятие. Мот. Навыки 99, 947–958.
Опубликовано Резюме | Опубликован полный текст
Stroop, JR (1935). Исследования вмешательства в серийных словесных реакций. Дж. Экспл. Психол . 18, 643–662.
Типпер, С. (1985). Отрицательный эффект прайминга: Ингибирующее праймирование игнорируемыми объектами. QJ Exp. Психол. А 37, 571–590.
Опубликовано Резюме | Опубликован полный текст
Ансуорт, Н., Хайц, Р. П., Шрок, Дж. К., и Энгл, Р. В. (2005). Автоматизированная версия задачи диапазона операций. Поведение. Рез. Методы 37, 498–505.
Опубликовано Резюме | Опубликован полный текст
Ансуорт, Н., Макмиллан, Б.Д., Брюэр, Г.А., и Спиллерс, Г.Дж. (2012). Ежедневные нарушения внимания: исследование индивидуальных различий. Дж. Экспл.Психол. Учиться. Мем. Код . 38, 1765–1772 гг.
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Велфорд, А. Т. (1958). Старение и человеческие способности . Лондон: Издательство Оксфордского университета.
Йи, Д. Дж., Вудман, Г. Ф., Виддерс, Д., Маруа, Р., и Чун, М. М. (2004). Нейронная судьба проигнорированных стимулов: диссоциирующие эффекты нагрузки на перцептивную и рабочую память. Нац. Нейроски . 7, 992–996.
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Янг, А.В., Эллис, А.В., Флуде, Б.М., Маквини, К.Х., и Хей, округ Колумбия (1986). Вмешательство «лицо-имя». Дж. Экспл. Психол. Гум. Восприятие. Выполните . 12, 466–475.
Опубликовано Резюме | Опубликован полный текст | Полнотекстовая перекрестная ссылка
Не думать о боли: роль дорсолатеральной префронтальной коры в модуляции боли | Мозг
Аннотация
Активность лобных долей во время боли обычно связана с обработкой внимания. Мы рассмотрели вопрос о том, диссоциируют ли анатомически в лобной доле обработку «снизу вверх» и модуляцию «сверху вниз» ноцицептивной информации с помощью ПЭТ-сканирования во время болезненной термической стимуляции нормальной кожи и кожи, обработанной капсаицином.Недавно мы показали, что боль после обычно безболезненных тепловых раздражителей на химически раздраженной коже (тепловая аллодиния) уникальным образом затрагивает обширные области двусторонней дорсолатеральной префронтальной (DLPFC), вентральной/орбитофронтальной (VOFC) и перигенуальной передней поясной извилины (ACC). Здесь мы применили анализ основных компонентов (PCA) и множественный регрессионный анализ для изучения ковариационной структуры интересующих объемов (VOI), активированных специально во время тепловой аллодинии у 14 здоровых мужчин, и оценили взаимосвязь этих VOI с оценками интенсивности боли и оказывать воздействие.В результате был получен первичный главный компонент (ПК), который положительно коррелировал с интенсивностью и неприятностью и объяснял активность в медиальном таламусе, билатеральном переднем островке, вентральном стриатуме, перигенуальной ППК и билатеральном VOFC. Действия в правом и левом DLPFC загружались на разные ПК и отрицательно коррелировали с воспринимаемой интенсивностью и неприятностью. Межрегиональная корреляция активности среднего мозга и медиального таламуса была значительно снижена во время высокой активности левой ДЛПФК, что позволяет предположить, что ее отрицательная корреляция с болевым аффектом может быть результатом ослабления эффективной связи между средним мозгом и медиальным таламическим путем.Напротив, активность правой ДЛПФК была связана с ослаблением связи передней доли островка как с интенсивностью боли, так и с аффектом. Мы предполагаем, что DLPFC осуществляет активный контроль над восприятием боли, модулируя корково-подкорковые и корково-кортикальные пути.
Введение
ЦНС способна изменять чувствительность к болевым раздражителям. Считается, что эндогенное торможение боли объясняет значительные колебания боли, возникающие в течение очень коротких периодов времени.Биологическое значение эндогенного контроля боли обычно рассматривается в контексте поведенческих конфликтов, когда индивидууму необходимо отключиться от боли, чтобы бороться или убежать при наличии телесных повреждений (Melzack and Casey, 1968). Аналогичными жизненными ситуациями человека являются спортивные соревнования и бои, во время которых субъект может не осознавать даже серьезных повреждений тканей, которые становятся болезненными, когда жертва прекращает заниматься этими действиями. Принимая во внимание, что спинальные и медуллярные механизмы тормозного контроля ноцицептивной передачи были в центре внимания обширных исследований, начиная с новаторских работ Мелзака и Уолла (1965), Басбаума и Филдса (1978) и Ле Барса и др. .(1979), у нас есть неполное понимание того, как высшие корковые функции способствуют эндогенному контролю боли. Поскольку боль трудно игнорировать и она мешает параллельной деятельности (Lorenz and Bromm, 1997; Eccleston and Crombez, 1999; Casey and Lorenz, 2000), вовлечение высших корковых функций может служить способом разрешения когнитивных и поведенческих конфликтов, позволяя конкурировать друг с другом. релевантные для задачи стимулы доминируют над болью.
Вероятной областью мозга-кандидатом для координации модуляции боли с целенаправленным поведением является лобная доля.Имеющиеся данные свидетельствуют о том, что дорсолатеральная префронтальная кора, включающая поля Бродмана 9 и 46, важна для постоянного наблюдения за внешним миром, сохранения информации в кратковременной памяти и управления эффективным контролем производительности в присутствии мешающих раздражителей (MacDonald et al ). ., 2000; Bunge et al. ., 2001; Sakai et al. ., 2002). Кроме того, электрическая стимуляция соединений волокон префронтальной коры со средним мозгом опосредует антиноцицептивные эффекты у грызунов (Cooper, 1975; Hardy and Haigler, 1985; Zhang et al ., 1998). Тем не менее, лобная доля может не играть единой роли в обработке боли, так как орбитофронтальные и медиальные лобные поражения уменьшают связанное с болью поведение у животных (Reshetniak and Kukushkin, 1989; Pastoriza et al ., 1996).
Неинвазивные нейровизуализационные исследования с использованием ПЭТ и функциональной МРТ позволяют нам изучить участие лобной доли в восприятии боли человеком. Различные группы описывают активность префронтальной коры после эксперимента (Casey et al ., 1996; Iadorola и др. ., 1998; Paulson и др. ., 1998; Baron и др. ., 1999; Tölle et al. ., 1999) или клинические (Hsieh et al. ., 1996; Rosen et al. ., 1996; Silverman et al. ., 1997) болевые состояния. Активность лобных долей во время боли обычно связана с когнитивной обработкой болевых раздражителей и вниманием (Coghill et al , 1999; Casey, 1999; Peyron et al , 1999; Bornhövd et al )., 2002). Имеются данные о том, что медиальные префронтальные области и перигенуальная передняя поясная кора (ACC) активируются ожиданием боли (Ploghaus et al. ., 1999; Sawamoto et al. ., 2000), взаимодействием боли с тревогой (Ploghaus et al. ., 1999). et al. ., 2001), плацебо-когниции (Petrovic et al. ., 2002) и когнитивные задачи (Petrovic et al. ., 2000; Bantick et al. ., 2002).
Недавно мы обнаружили существенную активацию префронтальной коры, включающую двустороннюю активность орбитофронтальной, перигенуальной поясной извилины и дорсолатеральной префронтальной коры, с помощью ПЭТ во время тепловых раздражителей на коже, обработанной капсаицином (Lorenz et al ., 2002). В этом исследовании мы сравнили одинаково интенсивные боли, адаптировав интенсивность теплового контактного датчика, применяемого к нормальной коже, и той же коже, чувствительной к капсаицину местного действия. Несмотря на равенство воспринимаемой интенсивности, вычитание изображения выявило уникальную активность медиально-таламического пути среднего мозга к лобной доле во время боли после обычно теплых раздражителей, воздействующих на сенсибилизированную кожу (тепловая аллодиния). Обычно мы интерпретировали сильную лобную активность во время тепловой аллодинии как отражение уникальных когнитивных и эмоциональных реакций на ноцицептивную информацию от патологической ткани.В настоящей работе мы использовали данные ПЭТ из нашего предыдущего исследования, чтобы определить, играют ли наблюдаемые активности в дорсолатеральной префронтальной, орбитофронтальной и перигенуальной областях ППК разные роли в обработке боли. Мы сосредоточились на оценке межрегиональной ковариационной структуры активности в интересующих объемах (VOI), активированных во время индуцированной капсаицином тепловой аллодинии, с использованием метода анализа основных компонентов (PCA). Кроме того, мы использовали регрессионный анализ, чтобы специально проверить гипотезу о том, что дорсолатеральная префронтальная кора (ДЛПФК) модулирует эффективную связь вдоль среднего мозга-таламического афферентного пути.
Методы
В исследовании приняли участие четырнадцать мужчин-правшей (средний возраст 23,9 ± 4,6 года ± стандартное отклонение). Местные институциональные наблюдательные советы Управления по делам ветеранов Анн-Арбора и Медицинских центров Мичиганского университета одобрили протокол до начала исследования. Психофизическая модель тепловой аллодинии, которая привела к уникальному включению активности лобных долей во время ПЭТ-сканирования H 2 15 O, подробно описана в нашей предыдущей статье (Lorenz et al ., 2002). Здесь мы ограничиваем описание теми аспектами протокола стимуляции и ПЭТ, которые важны для гипотезы настоящей статьи.
Капсаицин-тепловой стимул и психофизика
Мы доставляли тепловые стимулы к левому ладонному предплечью с помощью контактного термодатчика с обратной связью с цифровым управлением (Cygnus, Патерсон, Нью-Джерси, США), который имел площадь контакта с позолоченной медной поверхностью 254 мм 2 , нагреваемую постоянным током. Порядок сканирований и последовательность тестов и процедур внутри сканера приведены в таблице 1.Мы применяли одно состояние покоя без стимуляции (отсутствие контакта зонда) и два повторения двух температурно-временных профилей на коже левого предплечья. Мы выполнили первые пять сканирований до и последние пять сканирований после обработки кожи в течение 30 минут 1% капсаицином для местного применения (фильтровальная бумага размером 5 × 5 см, пропитанная 8-метил- N -ванилил-6-ноненамидом; Sigma, Сент-Луис, штат Миссури, США, разбавленный 70% этиловым спиртом). Индивидуальный порог тепловой боли определяли после помещения субъекта внутрь сканера.Тепловой зонд удерживался на левом предплечье испытуемого. Стимул начинался при 30 ° C и увеличивался со скоростью 0,9 ° C / с, пока субъект не отпускал кнопку, указывающую на начало боли. Температура в этот момент была отмечена, и среднее из пяти прогонов было принято за нормальный тепловой болевой порог (HPT n ). Мы измерили порог тепловой боли на сенсибилизированной коже (HPT c ) таким же образом после сканирования в покое после местного лечения капсаицином и после последнего сканирования.
Во время сканирования стимула датчик располагался на коже во время инъекции радиофармпрепарата и оставался неподвижным в течение 60 с сканирования, чтобы избежать механической стимуляции. Тепловой стимул начинался в начале сбора данных ПЭТ, примерно через 10 с после инъекции (см. ниже), снова с использованием времени нарастания 0,9 °C/с. Плато было либо на 2°C ниже, либо на 2°C выше HPT n . Место стимулируемой кожи перемещали между сканами против часовой стрелки в пределах площади 5 × 5 см.Легкая боль и тепло от одного капсаицина уменьшились в течение 20–30 минут после удаления пластыря. Таким образом, в то время как некоторый затяжной эффект самого капсаицина присутствовал во время сканирования в состоянии покоя после лечения, все последующие сканирования с тепловой стимуляцией происходили, когда это ощущение почти или полностью исчезало. Как было определено непрерывными оценками в пилотных тестах вне сканера (подробнее см. Lorenz et al ., 2002), низкоинтенсивный стимул ощущался как теплый на нормальной коже, тогда как лечение капсаицином делало его таким же болезненным, как и высокоинтенсивный. Интенсивный стимул на необработанной коже.Воспринимаемая интенсивность (0 = отсутствие ощущений, 10 = максимально болезненные) и неприятные ощущения (0 = не неприятные, 10 = максимально неприятные) сообщались в конце стимула после каждого сканирования с использованием устройства электронной визуальной аналоговой шкалы (ВАШ). Устройство состояло из горизонтального массива из 20 красных огней, которые показывались испытуемому, манипулируя потенциометром с ползунком. Экспериментатор считывал значение (напряжение) на цифровом дисплее сбоку устройства, невидимом для испытуемого.Маркер на этом устройстве указывал порог боли, который служил якорем для оценки интенсивности. После ознакомления с диапазоном интенсивностей стимулов испытуемые располагали маркер при определенном освещении, чтобы обозначить границу между предболевым и болевым диапазоном. Таким образом, ВАШ при нормальном пороге тепловой боли не был идентичен для всех субъектов и в среднем составлял 4,6 ± 0,85 (SD).
Для измерения качественных аспектов стимула мы использовали короткую форму опросника боли Макгилла (Melzack, 1987) после каждого сканирования стимуляции.Эта форма содержит 15 дескрипторов боли. Испытуемые отвечали «нет», «легкая», «умеренная» или «тяжелая», чтобы указать, насколько правильно слова описывают восприятие. Мы преобразовали ранжирование в числовые значения (0–3) и вычислили суммарные баллы отдельно для слов-дескрипторов сенсорного (например, горячий-обжигающий) и аффективного (например, наказание-жестокий) слов-дескрипторов.
ПЭТ
В этом исследовании использовался сканер Siemens/CTI 931/08-12 с 15 томографическими срезами, покрывающими аксиальное поле зрения 10 см.Для каждого из 10 сканирований каждый субъект получил болюсную инъекцию 50 мКи H 2 15 O в локтевую вену правой руки. Между каждым сканированием прошло не менее 15 минут. Сбор данных начался через 5 с после оценки поступления радиоактивности в мозг и продолжался в течение примерно 60 с. После нормализации каждого изображения, установленного для подсчета всего мозга (Fox and Raichle, 1984), изображения средней концентрации радиоактивности, оценивающие региональный мозговой кровоток (rCBF), были созданы для каждого экспериментального состояния у всех субъектов с помощью методов стереотаксической анатомической стандартизации (Minoshima et al ., 1994). Мы сделали изображения вычитания для каждого субъекта в соответствии с нашими экспериментальными переменными (отдых, две интенсивности, два состояния кожи). Мы провели повоксельный анализ статистического вычитания (карта оценок Z ) с поправкой на множественные сравнения на основе количества вокселей и гладкости случайной гауссовой модели после трехмерной фильтрации для увеличения соотношения сигнал-шум. соотношение и компенсировать анатомическую изменчивость (Friston et al ., 1991). Пространственное разрешение составляло 9 мм в соответствии с полной шириной на половине максимума функции Гаусса.Мы определили воксели со значительно увеличенным rCBF по сравнению со средней дисперсией шума, вычисленной для всех вокселей (объединенная дисперсия). Критический уровень значимости был определен путем корректировки ошибки типа I ( Z = 4,0, P < 0,05) с использованием этой информации, которая соответствует нескорректированному значению P , равному 0,0001 (Worsley et al ., 1992). ).
Взаимосвязь между психофизикой и реакцией кровотока
Используя метод, описанный ранее (Casey et al ., 1996), мы разработали VOI из изображения вычитания, на котором сравнивали низкоинтенсивный стимул на обработанной капсаицином коже (тепловая аллодиния) с высокоинтенсивным стимулом на нормальной коже (тепловая боль). Мы начали с вокселей, показывающих значительное пиковое увеличение rCBF в этом сравнении ( Z > 3,5), и постепенно увеличивали объем в трех измерениях, чтобы включить только те непрерывные воксели с увеличением rCBF, которое было значительно больше, чем глобальное среднее изменение ( P < 0.05, без поправок на множественные сравнения). Мы рассчитали коэффициенты корреляции ( r ) между активностью VOI и субъективными оценками отдельно для сканирования нормальной и сенсибилизированной кожи. В качестве первого шага мы удалили межиндивидуальную дисперсию, введя субъектов в качестве регрессоров, и сопоставили стандартизированную остаточную дисперсию с оценками интенсивности. Затем мы использовали остаточную дисперсию этого вычисления для расчета корреляции с оценками неприятности. Эта процедура устранила дисперсию, уникальную для каждого человека, но постоянную в экспериментальных условиях, и объяснила сильную связь между воспринимаемой интенсивностью и неприятностью.
PCA деятельности VOI
На другом этапе активности VOI, исключенные из межиндивидуальной дисперсии, подвергали PCA с варимаксным вращением, опять же отдельно для сканирования при нормальных и сенсибилизированных состояниях кожи. PCA обычно описывает наблюдаемые данные (здесь активность VOI) как взвешенную сумму статистически некоррелированных компонентов и вычисляет оценки основных компонентов (PC), которые количественно определяют, как каждый извлеченный компонент представляет данное измерение. Затем баллы ПК позволяют проводить дальнейший статистический анализ (здесь корреляция с оценками интенсивности и неприятности).Каждый компонент включает те измерения, которые подобны друг другу, т.е. дают высокую ковариацию. Количество ортогональных компонентов ограничено в соответствии с порогом значимости, определяемым тестом осыпи (Cattel, 1966), который ранжирует собственные значения собственных векторов компонентов, полученных из матрицы дисперсии-ковариации. В этом исследовании значимыми считались компоненты с собственными значениями >1. График факторной нагрузки показывает, как данная переменная представлена в каждом компоненте.Во время вращения варимакс оси извлеченных компонентов вращаются до тех пор, пока сумма квадратов факторных нагрузок не достигнет максимума. Эта процедура обычно характеризует более простую, более легко интерпретируемую структуру извлеченных компонентов только с высокими или низкими значениями загрузки по введенным переменным. Чтобы разумно свести к минимуму количество переменных, мы усреднили значения из вентролатеральной, вентромедиальной и орбитофронтальной VOI одного полушария вместе, впоследствии названного вентрально-орбитофронтальной корой (VOFC).Полученная матрица дисперсии-ковариации включала 10 VOI для общего количества 70 сканирований, что соответствует пяти испытаниям (одно в покое и два повторения низкой и высокой температуры стимула) у 14 субъектов при нормальном или сенсибилизированном состоянии кожи.
Наконец, мы проверили конкретную гипотезу о том, модулирует ли DLPFC связь между средним мозгом и медиальным таламусом. Мы выбрали эти две области мозга, потому что они представляют собой важный путь, связывающий спинной мозг с лимбической частью переднего мозга (Nauta and Kuypers, 1958; Melzack and Casey, 1968; Price, 2000).Основное предположение заключалось в том, что область (здесь DLPFC) модулирует величину коррелированной активности нейронов между двумя областями, указывая на силу потока нейронной информации между ними (Friston et al , 1995; Salinas and Sejnowski, 2001). Используя процедуру, описанную Бюхелем и Фристоном (1997), мы разделили 70 сканирований на две группы в соответствии со средним полураспределением высокой и низкой активности ДЛПФК и рассчитали корреляцию между активностью среднего мозга и медиального таламуса отдельно для сканирований с низкой и низкой активностью. высокая активность ДЛПФК.Мы количественно оценили статистическую значимость влияния активности ДЛПФК на корреляцию активности между этими двумя областями путем вычисления статистики F и значений Р для члена взаимодействия (средний мозг × ДЛПФК) в присутствии двух основных эффектов (средний мозг + DLPFC) в соответствии со следующей моделью линейной регрессии:
Таламус = b1 средний мозг + b2 DLPFC + b3 средний мозг × DLPFC
Хотя мы не смогли определить направление модулирующего действия, подразумеваемое термином взаимодействия, мы основываем нашу интерпретацию на нисходящей модели функции DLPFC при передаче афферентного сигнала, поддерживаемой нейрофизиологической фоновой информацией (см. Обсуждение).Регрессионный анализ и PCA были выполнены с использованием статистического пакета SPSS 10.0 (SPSS, Чикаго, Иллинойс, США).
Результаты
Психофизика и карты Z-счетов rCBF
Среднее значение оценки интенсивности по ВАШ подтверждает результаты пилотных испытаний о том, что низкоинтенсивный раздражитель на коже, обработанной капсаицином, индуцировал субъективную интенсивность, сходную с интенсивностью, вызываемой высокоинтенсивным раздражителем на нормальной коже ( t = 0,13, Р = 0.90; рис. 1, вверху). Хотя оценка неприятности этих состояний по ВАШ также не отличалась статистически (90 389 t 90 390 = 1,13, 90 389 P 90 390 = 0,28), неприятные ощущения, как правило, усиливались при лечении капсаицином в большей степени, чем интенсивность, о чем свидетельствует значимое взаимодействие состояния кожи и размера по ВАШ. ( F = 12,61, P < 0,01) в повторных измерениях ANOVA (дисперсионный анализ). Кроме того, во время низкоинтенсивной стимуляции кожи, обработанной капсаицином, наблюдался значительно более высокий эмоциональный балл по опроснику Макгилла по сравнению с высокоинтенсивной стимуляцией нормальной кожи ( t = 2.1, P = 0,05), тогда как сенсорная оценка не различалась ( t = 1,4, P = 0,17).
ПЭТ-изображения на рис. 1 показывают в левой и средней колонках карты Z -баллов, полученные путем вычитания сканов в нормальном состоянии покоя из сканов во время стимуляции высокоинтенсивным стимулом (HPT n + 2° C) на нормальной коже (например, тепловая боль) и при сканировании во время низкоинтенсивного стимула (HPT n – 2°C) на коже, обработанной капсаицином (т.е. тепловая аллодиния). Правый столбец нижней части рис. 1 показывает карту Z баллов, сравнивающую эти стимулы друг с другом. Четыре ряда относятся к разным уровням горизонтального среза относительно плоскости передней и задней комиссуры (уровень AC-PC, третий ряд). Несмотря на равенство воспринимаемой интенсивности, тепловая аллодиния приводила к значительному повышению активности в медиальном таламусе, билатеральном переднем островке, правом вентральном стриатуме, перигенуальной ACC)/медиальной префронтальной коре, билатеральных DLPFC и VOFC и дорсомедиальном отделе среднего мозга.Активации, которые присутствуют с одинаковой величиной в обоих условиях, удаляются путем вычитания; это включает ответы в ипсилатеральном вентролатеральном таламусе, SII/заднем островке, дорсальных частях правого полосатого тела и правой теменной нижней дольке (поле Бродмана 40). Поверхностные боковые и верхние проекции и парамедианные сагиттальные срезы на рис. 2 демонстрируют экстенсивное вовлечение VOFC и DLPFC обоих полушарий при тепловой аллодинии по сравнению с такой же интенсивной нормальной тепловой болью.Он также иллюстрирует заметную активность в вентральном стриатуме базальных ганглиев с участием прилежащего ядра правого полушария. Значительная дезактивация rCBF в этом сравнении проявилась в двусторонних язычных и веретенообразных извилинах затылочной доли и задней части поясной извилины (не показано). Эти дезактивации были связаны с высоким кровотоком в этих областях мозга в нормальном состоянии покоя, который был значительно более снижен во время тепловой аллодинии по сравнению с нормальной тепловой болью.Подобная деактивация зрительной системы наблюдалась в многочисленных исследованиях ПЭТ при различных стимулах и условиях выполнения задач и интерпретируется как неспецифический переход от режима активности мозга по умолчанию (отдых бодрствования) к умственной активности (Raichle et al ). ., 2001).
Анатомические координаты 17 VOI, которые превысили Z баллов 3,5 при сравнении тепловой аллодинии с одинаково интенсивной нормальной тепловой болью, и их корреляция с воспринимаемой болью и неприятными ощущениями, приведены в таблице 2.Активация только одной области, в правом VOFC ( x = 33, y = 55, z = 4), значительно коррелировала с воспринимаемой интенсивностью при стимуляции нормальной кожи. Однако при стимуляции сенсибилизированной кожи наблюдалась положительная корреляция с воспринимаемой интенсивностью в семи областях и с неприятными ощущениями в трех областях. rCBF в дорсомедиальном отделе среднего мозга, различных областях правой и левой VOFC и незначительно в медиальном таламусе коррелирует только с воспринимаемой интенсивностью.Левая и правая передняя островковая доля демонстрировали избирательную корреляцию с неприятными ощущениями. Одна область в левой ЛОК ( x = -42, y = 44, z = 4) показала положительную корреляцию как с воспринимаемой интенсивностью, так и с неприятными ощущениями. Хотя во время тепловой аллодинии активировалась перигенуальная АСС, не было корреляции ни с воспринимаемой интенсивностью, ни с оценкой неприятности. Примечательно, что левая DLPFC показала значительную отрицательную корреляцию с неприятными ощущениями и незначительную отрицательную корреляцию с воспринимаемой интенсивностью.Правый DLPFC также показал незначительно значимую отрицательную корреляцию с воспринимаемой интенсивностью ( P = 0,07) во время сканирования на чувствительной коже.
PCA
Мы выполнили PCA с поворотом варимакс на данных для 10 VOI, полученных из карты вычитания сканов с низкой интенсивностью на сенсибилизированной коже (тепловая аллодиния) за вычетом высокой интенсивности на нормальной коже (тепловая боль) отдельно для сканирований при нормальных и сенсибилизированных состояниях кожи. . Снижение до 10 по сравнению с исходными 17 VOI связано с усреднением различных субрегионов внутри VOFC (см. Методы).PCA извлек пять компонентов из сканов при нормальном состоянии кожи и три компонента из сканов при сенсибилизированном состоянии кожи. В таблице 3 перечислены собственные значения, проценты объясненной дисперсии и коэффициенты корреляции оценок факторов с воспринимаемой интенсивностью и неприятностью. Величина кумулятивно объясненной дисперсии этих компонентов составила 84 и 68% для нормальной и сенсибилизированной кожи соответственно. Примечательно, что только для сканирований во время состояния сенсибилизированной кожи показатели ПК давали связь с субъективными оценками, причем ПК 1 положительно, а ПК 2 отрицательно коррелировали как с оценками интенсивности, так и с оценками неприятности.ПК 3 имел уникальную отрицательную корреляцию с неприятностью.
График факторной нагрузки 10 VOI по трем параметрам PC, полученный на основе данных во время состояния сенсибилизированной кожи, представлен на рис. 3. Он отображает группу областей, которые положительно нагружаются на PC 1 и вовлекают медиальный таламус, правый вентральный стриатум, двусторонняя передняя островковая кора, перигенуальная ACC и двусторонняя VOFC. ПК 2 и ПК 3 в основном связаны с активностью правой и левой ДЛПФК соответственно.Средний мозг загружался отрицательно на ПК 2 и слабо положительно на ПК 1 и ПК 3. Таким образом, факторный анализ показал, что двусторонняя активность ДЛПФК варьировала независимо от активности в других областях, специфически вовлеченных во время тепловой аллодинии. Более того, регрессия психофизических данных показала обратную зависимость DLPFC от величины воспринимаемой боли. Правый и левый DLPFC имели разную факторную нагрузку и показали относительные различия в отношении корреляций с оценками интенсивности и неприятности.
Модуляция эффективной связи между средним мозгом и таламусом с помощью DLPFC
Чтобы проверить гипотезу о том, что ДЛПФК осуществляет нисходящую модуляцию эффективной связи вдоль среднего мозга-таламического пути, мы разделили 70 сканирований во время испытаний капсаицина на изображения с низкой и высокой активностью в ДЛПФК левого и правого полушарий по медианное полурасщепление данных VOI. На рис. 4 (вверху) показана обратная связь рейтинга неприятности с активностью левой ДЛПФК.VAS для неприятности был значительно выше при низкой по сравнению с высокой активностью DLPFC ( t = 4,25, P < 0,001). Нижняя диаграмма показывает, что корреляция активности среднего мозга и медиального таламуса также была значительно выше между сканированиями во время низкой активности левой ДЛПФК ( r = 0,72, P <0,001) по сравнению со сканированием во время высокой активности левой ДЛПФК ( r = 0,12, P = 0,49). Значение F взаимодействия левой ДЛПФК и среднего мозга на активность медиального таламуса было очень значимым ( F = 19.3, P < 0,0001). Сходный, но более слабый модулирующий эффект левой ДЛПФК наблюдался для корреляции между активностью среднего мозга и перигенной ППК ( F = 5,9, P = 0,02). Примечательно, что связь перигенуальной активности ACC с неприятными ощущениями также зависела от активности левой DLPFC. В то время как корреляция была положительной во время низкой активности левой ДЛПФК, она была отрицательной во время высокой активности левой ДЛПФК, что делало F значение взаимодействия (ДЛПФК × перигенуальная ACC) по ВАШ для неприятности значимой ( F = 4.9, P = 0,03). Этот эффект может объяснить, почему нам не удалось обнаружить корреляцию перигенуальной АКК с неприятными ощущениями, не зависящую от активности левой ДЛПФК.
Хотя правая ДЛПФК не смогла значительно изменить корреляцию между активностью среднего мозга и таламуса, взаимосвязь активности в передних островках обоих полушарий с интенсивностью боли и неприятными ощущениями поразительно зависела от величины активности правой ДЛПФК (рис. 5). Положительные корреляции наблюдались при низкой активности правой DLPFC, но не при высокой активности правой DLPFC.Статистика F для взаимодействия между правой ДЛПФК и правой передней островковой долей по ВАШ для интенсивности ( F = 9,7, P <0,01) и по ВАШ для неприятности ( F = 10,8; P <) был значительным. Точно так же наблюдалась значительная взаимосвязь между правым ДЛПФК и левым передним островком по ВАШ для интенсивности ( F = 5,9, P < 0,05) и ВАШ для неприятности ( F = 13,3, P = 0.001).
Обсуждение
Это исследование показывает, что наличие сенсибилизации ноцицепторов после местного лечения капсаицином значительно усиливает вовлечение лобных долей во время болезненной тепловой стимуляции. В нашей сопутствующей статье (Lorenz et al. ., 2002) мы подробно рассмотрели вопрос о том, что боль в ответ на обычно теплый раздражитель после лечения капсаицином (тепловая аллодиния) — это не просто сдвиг влево функции стимул-реакция тепловой реакции. ноцицепция.Вместо этого, по сравнению с такой же интенсивной нормальной болью от жара, аллодиния тепла включает относительно большее вовлечение медиальных таламических путей, которые передают более сильные аффективные реакции и, возможно, другую контекстуально важную качественную информацию, связанную с предполагаемой патологией ткани. Используя дополнительные статистические методы (PCA, регрессионный анализ), настоящее исследование демонстрирует диссоциацию субрегионов в лобной доле в отношении боли и модуляции боли. Сеть из 17 различных анатомических областей, специально активируемых во время тепловой аллодинии, показала, что двусторонние ДЛПФК исключительно отрицательно коррелируют с воспринимаемой интенсивностью и/или неприятностью.Правый и левый DLPFC учитывали уникальные факторы PCA матрицы дисперсии-ковариации при повторных сканированиях после лечения капсаицином, отличные от фактора PCA, который включал такие области, как медиальный таламус, перигенуальную ППК, правый вентральный стриатум, двусторонние островки и VOFC, и это положительно коррелировало с интенсивностью боли и неприятными ощущениями. Порегиональная корреляция активности между средним мозгом и медиальным таламусом, а также между средним мозгом и перигенной ППК была значительно выше при сканировании с низкой по сравнению с высокой активностью в левой ДЛПФК.Считается, что высокая корреляция указывает на усиленный поток нейронной информации, то есть на высокую эффективную связь (Салинас и Сейновски, 2001). Подход с использованием регрессионного анализа для выявления изменений в эффективной связности был описан для ПЭТ и функциональной МРТ зрительной системы (Friston et al. ., 1995; Büchel and Friston, 1997). Хотя направление модулирующего действия не может быть определено таким образом, наш результат может указывать на нисходящий способ ингибирования связи нейронов вдоль восходящего пути среднего мозга-таламо-поясной извилины через нисходящие волокна из префронтальной коры.Эта гипотеза согласуется с инвазивными исследованиями на животных. Hardy и Haigler (1985) продемонстрировали, что электрическая стимуляция префронтальной коры у крыс угнетает реакцию среднего мозга на вредные стимулы (ущипывание лапы). У кошек электрическая стимуляция околоводопроводного серого вещества или лобной (околокрестовой) коры подавляет ответ медиального таламуса на вредные раздражители (Andersen, 1986). Существование пути модуляции боли, включающего средний мозг грызунов, медиальный таламус и префронтальную кору, также подтверждается исследованиями Condes-Lara et al .(1989).
Наш вывод о том, что усиленная связь между средним мозгом и медиальным таламусом связана с повышенным рейтингом неприятности во время низкой активности левой ДЛПФК, согласуется с ролью этих областей в опосредовании аффективных реакций на боль (Nauta and Kuypers, 1958; Melzack and Casey, 1968). ; Прайс, 2000). Активность левой ДЛПФК была дополнительно связана со снижением корреляции перигенуальной АКК с воспринимаемой неприятностью. Перигенуальная ППК получает основной сигнал от медиального таламуса и тесно связана с прилежащим ядром вентрального полосатого тела (Kunishio and Haber, 1994), которое также заметно активировалось при стимуляции кожи, обработанной капсаицином.Эти две области мозга, по-видимому, представляют собой узлы перекрытия схем боли и вознаграждения (Becerra et al ., 2001). Интересно, что прилежащее ядро оказывает антиноцицептивное действие после ядовитого капсаицина и термических раздражителей (Gear et al. ., 1999). Напротив, правый DLPFC в первую очередь снижает корреляцию билатеральных передних островков как с воспринимаемой интенсивностью, так и с неприятными ощущениями. Островковая доля участвует в декодировании интенсивности тепловых раздражителей (Coghill et al ., 1999; Craig et al ., 2000) и получает сенсорную информацию от латеральных ядер таламуса (Craig and Dostrovsky, 1999). Отсутствие возможности наблюдать снижение эффективной связи между средним мозгом и таламусом во время высокой активности правой DLPFC может быть связано с тем, что мы ограничили наш анализ медиально-таламическим VOI, полученным из карты вычитания тепловой аллодинии за вычетом столь же интенсивной нормальной тепловой боли, которая заметно снижена активность латерального таламуса (рис. 1). Остается неясным, могут ли описанные различия между левой и правой ДЛПФК в PCA и регрессионном анализе быть связаны с межполушарной дифференцировкой механизмов модуляции префронтальной боли.
Поскольку основная цель этого исследования заключалась в том, чтобы продемонстрировать, что аллодиния тепла задействует супраспинальную сеть, отличную от той, которая задействована при такой же интенсивной нормальной боли, наши инструкции, данные испытуемым, не предназначались для манипулирования восприятием боли. Тем не менее, можно предположить, что испытуемые лучше когнитивно контролировали боль во время стимуляции обработанной капсаицином кожи, которую они воспринимали как раздраженную и более чувствительную. Гипноз избирательно изменяет аффективные реакции на боль в соответствии с изменениями в орбитофронтальной и средней поясной областях (Rainville et al ., 1997). Аналогичным образом, умственная нагрузка снижает восприятие боли, что согласуется со снижением активности в таламусе, островковой и средней части поясной извилины, а также с повышенной активностью в перигенуальной поясной извилине и лобной коре (Bantick et al ., 2002). Активность в этих последних областях мозга также усиливается и более сильно коррелирует с активностью среднего мозга во время плацебо-анальгезии (Petrovic et al ., 2002). Мы наблюдали более сильную корреляцию активности среднего мозга и перигенальной ППК во время сканирований с низкой активностью левой ДЛПФК, во время которых степень связи между перигенальной ППК и неприятными ощущениями была значительно выше, чем при сканировании с высокой активностью левой ДЛПФК.Возможно, что функциональная диссоциация в среднем мозге может объяснить как облегчающее, так и тормозящее влияние на боль и аффект (Hirakawa et al ., 2000).
Активность в VOFC отличалась от таковой в DLPFC тем, что в нашем исследовании была положительно коррелирована с болью. Это наблюдение согласуется с сообщениями об активности в этих областях во время ожидания боли (Ploghaus et al , 1999; Sawamoto et al , 2000) или ее обострения тревогой (Ploghaus et al )., 2001). Кроме того, у животных с орбитофронтальными (Reshetniak and Kukushkin, 1989) и медиально-лобными (Pastoriza et al ., 1996) поражениями болевой синдром снижен. Функциональная диссоциация вентромедиальной/орбитальной и латеральной префронтальной коры также была описана в исследованиях патологических биохимических процессов в мозге у пациентов с хронической болью (Grachev et al , 2000, 2002), дифференциальных репрезентаций отрицательных и положительных эмоций (Northoff et al ., 2000), а также сдвиги аффективных и когнитивно-внимательных настроек реакции (Dias et al ., 1996). Примечательно, что целевые области нисходящих проекций от лобной доли к среднему мозгу анатомически диссоциируют у крыс в отношении медиального или латерального происхождения в префронтальной коре (Hardy, 1986). Роль DLPFC в контроле боли, кроме того, согласуется с исследованиями, которые демонстрируют, что эта область связана с эффективной работой в присутствии противоречивых стимулов (MacDonald et al ., 2000; Bunge et al ., 2001). Задачи рабочей памяти, которые требуют высокой степени исполнительного контроля, обычно связаны с активностью DLPFC (Smith and Jonides, 1999; Funahashi, 2001).DLPFC может защищать поддержание сиюминутных поведенческих целей, делая операции рабочей памяти устойчивыми к отвлекающим стимулам (Sakai et al , 2002). Лоренц и Бромм (1997) обнаружили, что способность поддерживать точность выполнения задания на кратковременную память во время экспериментальной болезненной мышечной ишемии положительно коррелирует с активностью лобных долей, измеренной с помощью связанных с событиями мозговых потенциалов, вызванных стимулами памяти. Взаимодействие префронтальной коры со средним мозгом, таламическими, полосатыми и поясными структурами лимбической системы может, таким образом, отражать активное манипулирование поведенческим доминированием боли в зависимости от мотивационного и эмоционального контекста.Наличие аномального состояния ткани, такого как воспаление, может сильно задействовать префронтальные механизмы модуляции боли, потому что это требует поведенческой гибкости и способности подавлять доминантные тенденции реакции, чтобы гарантировать оптимальную адаптацию. Примечательно, что нисходящее торможение более эффективно задействуется при остром воспалении сустава грызунов по сравнению с хроническим, даже когда ноцицептивный тестовый стимул вызывает одинаковую интенсивность болевых реакций в любом состоянии (Danziger et al ., 1999). Поэтому возможно, что когнитивные механизмы контроля боли, управляемые префронтальной корой, синергетически взаимодействуют с автономными механизмами нисходящего торможения, управляемыми подкорковыми структурами, во время острых стадий травмы и воспаления. Напротив, острая боль от нормальной ткани требует менее гибкой, более привычной реакции на угрозу надвигающегося повреждения ткани. Активность, возникающая в более быстро проводящих Aδ-ноцицепторах, и проекция в пределах латерального таламического пути к ассоциативным областям сенсорной и моторной коры, а также вовлечение дорсальных базальных ганглиев и мозжечка кажутся подходящими для достижения этой функции и могут объяснить, почему эти области мозга преимущественно активны. активируется острой тепловой болью на нормальной коже (Casey et al ., 2001).
Хотя результаты этого исследования совместимы с ключевой ролью DLPFC в корковых механизмах модуляции боли, эта интерпретация основана на корреляционном анализе. Мы не можем исключить возможность того, что условия нашего эксперимента по-разному влияли на эту область мозга посредством восходящих механизмов. Необходим интервенционный подход для дальнейшего тестирования связи между активностью DLPFC и подавлением боли. В будущих исследованиях можно было бы изучить эффекты взаимодействий между различной интенсивностью боли и различной степенью нагрузки на рабочую память или использовать транскраниальную магнитную стимуляцию для обратимого и избирательного вмешательства в префронтальную функцию во время болевой стимуляции.
Благодарности
Мы выражаем признательность за поддержку Донне Дж. Кросс, Памеле Э. Полсон, Томасу Дж. Морроу, Эндрю Уидену и Эду Маккенне, а также благодарим Кристиана Бюхеля и Рольфа-Детлефа Триде за полезные предложения по предварительным версиям рукописи. Эта работа была поддержана Департаментом по делам ветеранов, грантом NIH Службы общественного здравоохранения США P01HD33986 и Министерством энергетики (DE-FG02-87-ER60561). JL был поддержан грантом от Фонда Макса Кейда.
Рис. 1 Регионарная мозговая активность во время одинаково интенсивной боли при нормальных состояниях кожи и при лечении капсаицином. Стимуляция нормальной кожи с высокой интенсивностью вызывала ту же интенсивность боли, что и стимуляция низкой интенсивности на коже, обработанной капсаицином (вверху). Однако ПЭТ-изображения H 2 15 O во время тепловой боли (нижняя часть рисунка, левый столбец) и столь же интенсивной тепловой аллодинии (средний столбец) отличались по сравнению с нормальным состоянием покоя.Аналогичные величины активности в дорсальном стриатуме, латеральном таламусе (lat tha) и задней части островка (post ins) удаляются при вычитании изображения тепловой аллодинии минус тепловая боль (правая колонка), контрастной активности в вентральном стриатуме, медиальном таламусе (med tha), передняя островковая доля (ant ins), средний мозг, DLPFC, медиальная префронтальная кора, VOFC и перигенуальная ACC во время тепловой аллодинии.
Рис. 1 Регионарная мозговая активность во время одинаково интенсивной боли при нормальных состояниях кожи и при лечении капсаицином.Стимуляция нормальной кожи с высокой интенсивностью вызывала ту же интенсивность боли, что и стимуляция низкой интенсивности на коже, обработанной капсаицином (вверху). Однако ПЭТ-изображения H 2 15 O во время тепловой боли (нижняя часть рисунка, левый столбец) и столь же интенсивной тепловой аллодинии (средний столбец) отличались по сравнению с нормальным состоянием покоя. Аналогичные величины активности в дорсальном стриатуме, латеральном таламусе (lat tha) и задней части островка (post ins) удаляются при вычитании изображения тепловой аллодинии минус тепловая боль (правая колонка), контрастной активности в вентральном стриатуме, медиальном таламусе (med tha), передняя островковая доля (ant ins), средний мозг, DLPFC, медиальная префронтальная кора, VOFC и перигенуальная ACC во время тепловой аллодинии.
Рис. 2 Поверхностные изображения тепловой аллодинии за вычетом столь же интенсивной нормальной тепловой боли. Обратите внимание на широкое вовлечение активности лобных долей в области VOFC и DLPFC и заметную активность вентрального стриатума базальных ганглиев с участием прилежащего ядра. LAT = боковой; МЭД = медиальный; SUP = превосходный; dm = дорсомедиальный.
Рис. 2 Поверхностные изображения тепловой аллодинии за вычетом столь же интенсивной нормальной тепловой боли. Обратите внимание на широкое вовлечение активности лобных долей в области VOFC и DLPFC и заметную активность вентрального стриатума базальных ганглиев с участием прилежащего ядра.LAT = боковой; МЭД = медиальный; SUP = превосходный; dm = дорсомедиальный.
Рис. 3 Графики факторной нагрузки, полученные на основе активности PCA с вращением варимакс при состоянии сенсибилизированной кожи в 10 VOI, специфически активированных тепловой аллодинией. Обратите внимание, что активность DLPFC обоих полушарий нагружает селективный главный компонент (PC), отличный от PCA 1, что объясняет активность в областях мозга 2–8. m Tha = медиальный таламус; perig ACC = перигенуальный ACC; r v = правый вентральный; r ant INS = передний правый островок; l ant INS = левый передний островок; р = правильно; л = слева.
Рис. 3 Графики факторной нагрузки, полученные из варимакс-вращающегося PCA активности при сенсибилизированном состоянии кожи в 10 VOI, специфически активированном тепловой аллодинией. Обратите внимание, что активность DLPFC обоих полушарий нагружает селективный главный компонент (PC), отличный от PCA 1, что объясняет активность в областях мозга 2–8. m Tha = медиальный таламус; perig ACC = перигенуальный ACC; r v = правый вентральный; r ant INS = передний правый островок; l ant INS = левый передний островок; р = правильно; л = слева.
Рис. 4 Регрессия оценок неприятности по активности VOI в левой ДЛПФК (вверху). Обратите внимание на обратную связь болевого аффекта с активностью левой ДЛПФК ( r = –0,30, P <0,01) во время испытаний ( n = 70) на сенсибилизированной коже. Медиана половинного разделения данных показала значительно большую неприятность при низкой (светлые кружки), чем при высокой (закрашенные кружки) активности DLPFC. Зависимость корреляции активности между средним мозгом и медиальным таламусом от активности левой ДЛПФК (внизу).Выявлена значительно более низкая корреляция активности в среднем мозге с таковой в медиальном таламусе при высокой (закрашенные кружки; черная линия регрессии) активности по сравнению с низкой (светлые кружки; серая линия регрессии) активностью левой ДЛПФК.
Рис. 4 Регрессия оценок неприятности по активности VOI в левой ДЛПФК (вверху). Обратите внимание на обратную связь болевого аффекта с активностью левой ДЛПФК ( r = –0,30, P <0,01) во время испытаний ( n = 70) на сенсибилизированной коже.Медиана половинного разделения данных показала значительно большую неприятность при низкой (светлые кружки), чем при высокой (закрашенные кружки) активности DLPFC. Зависимость корреляции активности между средним мозгом и медиальным таламусом от активности левой ДЛПФК (внизу). Выявлена значительно более низкая корреляция активности в среднем мозге с таковой в медиальном таламусе при высокой (закрашенные кружки; черная линия регрессии) активности по сравнению с низкой (светлые кружки; серая линия регрессии) активностью левой ДЛПФК.
Рис.5 Корреляция активности в левой и правой передней доле островка с показателями интенсивности (вверху) и неприятности (внизу) по ВАШ при низкой (светлые кружки) и высокой (темные кружки) активности правой ДЛПФК.
Рис. 5 Корреляция активности в левом и правом переднем островке с оценками интенсивности (вверху) и неприятности (внизу) по ВАШ при низкой (светлые кружки) и высокой (темные кружки) активности правой ДЛПФК.
Таблица 1Расписание событий в сканер домашних животных
| 9 | Группа | Группа B | |
| HPT N N 74 N | |||
| 0 | Отдых | Отдых | Отдых |
| 15 | HPT N — 2 ° C | — 2 ° C 73 HPT N + 2 ° C||
| 30 773 HPT N + 2 ° C | HPT N — 2 ° C | 9 | 4 73 HPT N + 2 ° C | — 2 ° C N — 2 ° C — 2 ° C
| 60774 — 2 ° C | HPT N + 2 ° C | + 2 ° C||
| 70-100 | 70-100 | 90-10074 | |
| 110 | Отдых | Отдых | |
| 115 | HPT C | HPT C | |
| 125 | 3 HPT N — 2 ° C N — 2 ° C N — 2 ° C + 2 ° C + 2 ° C + 2 ° C|||
| 140 | 73 HPT N + 2 ° CHPT N — 2 ° C | — 2 ° C15543 HPT N + 2 ° C | — 2 ° C — 2 ° C — 2 ° C — 2 ° C — 2 ° C|
| HPT N — 2 ° C | HPT N + 2 ° C | ||
| 175 C C C C C 4 C |
| Время (мин) | Группа | Group B |
| -10 | HPT N | |
| 0 | Отдых | Отдых |
| 15 | HPT N — 2 ° C | 73 HPT N + 2 ° C|
| 30 | 303 HPT N + 2 ° C | + 2 ° C — 2 ° C — 2 ° C — 2 ° C — 2 ° C — 2 ° C9|
| 45 | 73 HPT N + 2 ° C — 2 ° C||
| 60772 | 60774 | HPT N — 2 ° C | — 2 ° C + 2 ° C + 2 ° C + 2 ° C + 2 ° C70-100 | Отдых |
| 110 | Отдых 9074 | |
| 115 | HPT C 4 70050 C C 90721254 | HPT N — 2 ° C | — 2 ° C + 2 ° C + 2 ° C 79
| 140 N + 2 ° C | — 2 ° C N — 2 ° C — 2 ° C — 2 ° C — 2 ° C — 2 ° C||
| HPT + 2 ° C | — 2 ° C — 2 ° C — 2 ° C||
| HPT N – 2°С | HPT N + 2 ° C | + 2 ° C|
| 175 | HPT C C 4 HPT C C
Расписание мероприятий в сканер домашних животных
| Мин) | Группа | Group B | 9 | HPT 73 HPT N HPT N | 0 | REST | REST |
| 3 15 73 HPT N — 2 ° C | 73 HPT N + 2 ° C + 2 ° C + 2 ° C + 2 ° C 779|||||
| 304 73 HPT N + 2 ° C | — 2 ° C N — 2 ° C — 2 ° C|||||
| HPT N + 2 ° C | HPT N — 2 ° C | — 2 ° C — 2 ° C — 2 ° C — 2 ° C||||
| 60 3 HPT N — 2 ° C | — 2 ° C + 2 ° C|||||
| 70-100 | Capsai Лечение CIN | ||||
| 110 | Остальной | Остальной | |||
| 115 | ТВД с | ТВД с | |||
| 125 | ТВД л — 2 ° С | ТВД n + 2 ° C | |||
| 140 | 73 HPT N + 2 ° C + 2 ° C — 2 ° C N — 2 ° C — 2 ° C9|||||
| 155 | 73 HPT N + 2 ° CHPT N — 2 ° C | — 2 ° C||||
| 1703 HPT N — 2 ° C | — 2 ° C + 2 ° C + 2 ° C 99|||||
| 175 | HPT C | 73 HPT C
Множественный регрессионный анализ rCBF в VOI, полученный в результате вычитания изображения сканирования во время низкоинтенсивной стимуляции на чувствительной коже (тепловая аллодиния) минус высокоинтенсивная стимуляция на нормальной коже (тепловая боль)
| Область | Стереотаксические координаты: | интенсивности боли | Pain неприятности | |||||||||||||||
| х , у , г (мм)) * | Нормального | Сенсибилизированные кожи | Нормальный | Сенсибилизированные кожи | ||||||||||||||
| г | т | Р | г | т | Р | г | т | Р | р | т | П 9077 4 | |||||||
| Dm средний мозг | 1, –33, –14 | 0.01 | 0,11 0,91 | 0,35 | 3,05 | <0.01 | -0,09 -0,74 | 0,46 -0,02 -0,16 | ||||||||||
| Дм Thalamus | 1, -17, 11 | , 11 | 0,08 | 0.62 | 0.54 | 0.54 | 0.08 | 0.09 | 0,09 | -0,13 | -1.03 | 0.31 | -0.02 | -0.02-0.14 | 0,89 | 9 | 9 | |
| R V Striatum | 17, 5, -2 | 3 -0.01 | -0,07 | 0.08 | 0.08 | 0.08 | 0.50 | 0,08 | 0.66 | 0.51 | 0,11 | 0,11 | 0.92 | 0.36 | 0.36 | |||
| l Муравей INSULA | -24, 10, 7 | -0-02 | -0.22 | 3 0.83 | 0,17 | 1,38 | 0,17 | 0,18 | 1,45 | 0,14 | 0,37 | 3,29 | <0.01 | |||||
| R муравей островковой | 42, 5, 11 | 0.00 | 0,00 | 0,00 | 1.0 | 0,02 | 0,19 | 0,85 | 0,85 | 0,84 | 0,41 | 0,41 | 0,25 2.16 | 0,03 | ||||
| Perig АСС | -1, 37, 16 | -0,04 -0,36 | 0,72 0,08 | 0,65 | 0,52 | -0,04 -0,34 | 0.73 | -0774 | -0.01 | -0.11 | 0,11 | |||||||
| R VOFC | R VOFC | 12, 44, 2 | 0,01 | 0,09 | 0.09 | 0,22 | 3 1.86 | 0,07 | 0,00 | 0,00 | 1,0 | 0,15 | 1,23 | 0,22 | ||||
| 17, 57, 4 | 0,03 | 0,25 | 0,80 | 0,29 | 2.50 | 4 | 0.02 0.02 4 | -0.13 | -1.10 | 0.28 | 0,28 | 0,19 | 1,55 | 0,13 | ||||
| 33, 55, 4 | 0.23 | 1,96 | 0,05 | 0,23 | 1,96 | 0,05 | 0,05 | 0,41 | 0,68 | 0,12 | 0,97 | 0,33 | ||||||
| 28, 53, -2 | -0774 | -0.04-0.32 | 0.75 | 0,75 | 0,17 | 3 1.39 | 3 0,17 | 9074 9073 -0,04-0.31 | 0,76 -0,03 -0,21 | |||||||||
| 35, 46, 18 | 0,03 | 0,24 | 0,81 | -0,06 -0,51 | 0,62 -0,14 | -1.14 | 3 -1.14 | 0,26 | -0.07 | -0.62 | 0.54 | 4 | ||||||
| 28, 46, -4 | -0.09 | — 0,76 | 3 0,45 | 0.24 | 2,04 | 0,05 | 0,15 | 1,23 | 0,22 | 0,22 | 1,89 | 0,06 | ||||||
| л VOFC | -28, 50, 7 | 0,07 | 0,55 | 0.58 | 0.33 | 2 | 3 2,90 4 | <0,01 | -0.01 | -0 0.93 | 0.93 | 3 0,11 | 3 0.87 | 0,39 | | ||||
| -42, 44, | 4 0.12 | 1,00 0,32 | 0,26 | 2,21 | 0,03 | 0,08 | 0,65 | 0,52 | 0.27 2 232 | 0,02 | ||||||||
| -21, 46, -7 | -7 | 9074-0.29 | 0.77 | 0,35 | 3,04 | <0.01 | 0,07 | 0,56 | 0,58 | 0,20 | 1,7 | 0,10 | ||||||
| R ДЛПФК | 33, 14, 45 | -0.13 | -1.05 | -1.050.30 | -0.22 | -1.84 | 0,07 | -0,13 | -1 09 | — 0.30 | -0,19 | 3 -1.61 | 0,11 | |||||
| л ДЛПФК | -37, 17, 36 | -0,01 -0,09 | 0,93 -0,13 -1,07 | 0,29 -0,15 -1,21 | 0,23 | -0,30 -2,62 | 0,01 | |||||||||||
| область Стереотаксические координаты: | интенсивности боли | Pain неприятности | ||||||||||||||||
| х, у , г (мм)) * | Нормальных | Сенсибилизированные кожи | Нормальных | Сенсибилизированные кожи | ||||||||||||||
| г | т | Р | г | т | П | р | т | 9 0773 P PR 3 R | T | P | ||||||||
| DM Midbrain | 1, -33, -14 | 0.01 | 0,11 0,91 | 0,35 | 3,05 | <0.01 | -0,09 -0,74 | 0,46 -0,02 -0,16 | ||||||||||
| Дм Thalamus | 1, -17, 11 | , 11 | 0,08 | 0.62 | 0.54 | 0.54 | 0.08 | 0.09 | 0,09 | -0,13 | -1.03 | 0.31 | -0.02 | -0.02-0.14 | 0,89 | 9 | 9 | |
| R V Striatum | 17, 5, -2 | 3 -0.01 | -0,07 | 0.08 | 0.08 | 0.08 | 0.50 | 0,08 | 0.66 | 0.51 | 0,11 | 0,11 | 0.92 | 0.36 | 0.36 | |||
| l Муравей INSULA | -24, 10, 7 | -0-02 | -0.22 | 3 0.83 | 0,17 | 1,38 | 0,17 | 0,18 | 1,45 | 0,14 | 0,37 | 3,29 | <0.01 | |||||
| R муравей островковой | 42, 5, 11 | 0.00 | 0,00 | 0,00 | 1.0 | 0,02 | 0,19 | 0,85 | 0,85 | 0,84 | 0,41 | 0,41 | 0,25 2.16 | 0,03 | ||||
| Perig АСС | -1, 37, 16 | -0,04 -0,36 | 0,72 0,08 | 0,65 | 0,52 | -0,04 -0,34 | 0.73 | -0774 | -0.01 | -0.11 | 0,11 | |||||||
| R VOFC | R VOFC | 12, 44, 2 | 0,01 | 0,09 | 0.09 | 0,22 | 3 1.86 | 0,07 | 0,00 | 0,00 | 1,0 | 0,15 | 1,23 | 0,22 | ||||
| 17, 57, 4 | 0,03 | 0,25 | 0,80 | 0,29 | 2.50 | 4 | 0.02 0.02 4 | -0.13 | -1.10 | 0.28 | 0,28 | 0,19 | 1,55 | 0,13 | ||||
| 33, 55, 4 | 0.23 | 1,96 | 0,05 | 0,23 | 1,96 | 0,05 | 0,05 | 0,41 | 0,68 | 0,12 | 0,97 | 0,33 | ||||||
| 28, 53, -2 | -0774 | -0.04-0.32 | 0.75 | 0,75 | 0,17 | 3 1.39 | 3 0,17 | 9074 9073 -0,04-0.31 | 0,76 -0,03 -0,21 | |||||||||
| 35, 46, 18 | 0,03 | 0,24 | 0,81 | -0,06 -0,51 | 0,62 -0,14 | -1.14 | 3 -1.14 | 0,26 | -0.07 | -0.62 | 0.54 | 4 | ||||||
| 28, 46, -4 | -0.09 | — 0,76 | 3 0,45 | 0.24 | 2,04 | 0,05 | 0,15 | 1,23 | 0,22 | 0,22 | 1,89 | 0,06 | ||||||
| л VOFC | -28, 50, 7 | 0,07 | 0,55 | 0.58 | 0.33 | 2 | 3 2,90 4 | <0,01 | -0.01 | -0 0.93 | 0.93 | 3 0,11 | 3 0.87 | 0,39 | | ||||
| -42, 44, | 4 0.12 | 1,00 0,32 | 0,26 | 2,21 | 0,03 | 0,08 | 0,65 | 0,52 | 0.27 2 232 | 0,02 | ||||||||
| -21, 46, -7 | -7 | 9074-0.29 | 0.77 | 0,35 | 3,04 | <0.01 | 0,07 | 0,56 | 0,58 | 0,20 | 1,7 | 0,10 | ||||||
| R ДЛПФК | 33, 14, 45 | -0.13 | -1.05 | -1.050.30 | -0.22 | -1.84 | 0,07 | -0,13 | -1 09 | — 0.30 | -0,19 | 3 -1.61 | 0,11 | |||||
| л ДЛПФК | -37, 17, 36 | -0,01 -0,09 | 0,93 -0,13 -1,07 | 0,29 -0,15 -1,21 | 0.23 | -0774 | -0.30 | -2.62 | 3 -2.62 | 0,01 | 0,01 | |||||||
Многократный регрессионный анализ RCBF в Voi, полученный в результате вычитания изображений для сканов при низкой интенсивности на сенсибилизированную кожу (тепловысокая аллодиния) минус высокой интенсивности Стимуляция на нормальной коже (боли нагрева)
| регион | Нормальная | Чувствительная кожа | Нормальная | Чувствительная кожа | ||||||||||||||
| г | т | Р | г | т | Р | г | т | Р | г | t | P | |||||||
| Dm средний мозг | 1, –33, –14 | 0.01 | 0,11 0,91 | 0,35 | 3,05 | <0.01 | -0,09 -0,74 | 0,46 -0,02 -0,16 | ||||||||||
| Дм Thalamus | 1, -17, 11 | , 11 | 0,08 | 0.62 | 0.54 | 0.54 | 0.08 | 0.09 | 0,09 | -0,13 | -1.03 | 0.31 | -0.02 | -0.02-0.14 | 0,89 | 9 | 9 | |
| R V Striatum | 17, 5, -2 | 3 -0.01 | -0,07 | 0.08 | 0.08 | 0.08 | 0.50 | 0,08 | 0.66 | 0.51 | 0,11 | 0,11 | 0.92 | 0.36 | 0.36 | |||
| l Муравей INSULA | -24, 10, 7 | -0-02 | -0.22 | 3 0.83 | 0,17 | 1,38 | 0,17 | 0,18 | 1,45 | 0,14 | 0,37 | 3,29 | <0.01 | |||||
| R муравей островковой | 42, 5, 11 | 0.00 | 0,00 | 0,00 | 1.0 | 0,02 | 0,19 | 0,85 | 0,85 | 0,84 | 0,41 | 0,41 | 0,25 2.16 | 0,03 | ||||
| Perig АСС | -1, 37, 16 | -0,04 -0,36 | 0,72 0,08 | 0,65 | 0,52 | -0,04 -0,34 | 0.73 | -0774 | -0.01 | -0.11 | 0,11 | |||||||
| R VOFC | R VOFC | 12, 44, 2 | 0,01 | 0,09 | 0.09 | 0,22 | 3 1.86 | 0,07 | 0,00 | 0,00 | 1,0 | 0,15 | 1,23 | 0,22 | ||||
| 17, 57, 4 | 0,03 | 0,25 | 0,80 | 0,29 | 2.50 | 4 | 0.02 0.02 4 | -0.13 | -1.10 | 0.28 | 0,28 | 0,19 | 1,55 | 0,13 | ||||
| 33, 55, 4 | 0.23 | 1,96 | 0,05 | 0,23 | 1,96 | 0,05 | 0,05 | 0,41 | 0,68 | 0,12 | 0,97 | 0,33 | ||||||
| 28, 53, -2 | -0774 | -0.04-0.32 | 0.75 | 0,75 | 0,17 | 3 1.39 | 3 0,17 | 9074 9073 -0,04-0.31 | 0,76 -0,03 -0,21 | |||||||||
| 35, 46, 18 | 0,03 | 0,24 | 0,81 | -0,06 -0,51 | 0,62 -0,14 | -1.14 | 3 -1.14 | 0,26 | -0.07 | -0.62 | 0.54 | 4 | ||||||
| 28, 46, -4 | -0.09 | — 0,76 | 3 0,45 | 0.24 | 2,04 | 0,05 | 0,15 | 1,23 | 0,22 | 0,22 | 1,89 | 0,06 | ||||||
| л VOFC | -28, 50, 7 | 0,07 | 0,55 | 0.58 | 0.33 | 2 | 3 2,90 4 | <0,01 | -0.01 | -0 0.93 | 0.93 | 3 0,11 | 3 0.87 | 0,39 | | ||||
| -42, 44, | 4 0.12 | 1,00 0,32 | 0,26 | 2,21 | 0,03 | 0,08 | 0,65 | 0,52 | 0.27 2 232 | 0,02 | ||||||||
| -21, 46, -7 | -7 | 9074-0.29 | 0.77 | 0,35 | 3,04 | <0.01 | 0,07 | 0,56 | 0,58 | 0,20 | 1,7 | 0,10 | ||||||
| R ДЛПФК | 33, 14, 45 | -0.13 | -1.05 | -1.050.30 | -0.22 | -1.84 | 0,07 | -0,13 | -1 09 | — 0.30 | -0,19 | 3 -1.61 | 0,11 | |||||
| л ДЛПФК | -37, 17, 36 | -0,01 -0,09 | 0,93 -0,13 -1,07 | 0,29 -0,15 -1,21 | 0,23 | -0,30 -2,62 | 0,01 | |||||||||||
| область Стереотаксические координаты: | интенсивности боли | Pain неприятности | ||||||||||||||||
| х, у , г (мм)) * | Нормальных | Сенсибилизированные кожи | Нормальных | Сенсибилизированные кожи | ||||||||||||||
| г | т | Р | г | т | П | р | т | 9 0773 P PR 3 R | T | P | ||||||||
| DM Midbrain | 1, -33, -14 | 0.01 | 0,11 0,91 | 0,35 | 3,05 | <0.01 | -0,09 -0,74 | 0,46 -0,02 -0,16 | ||||||||||
| Дм Thalamus | 1, -17, 11 | , 11 | 0,08 | 0.62 | 0.54 | 0.54 | 0.08 | 0.09 | 0,09 | -0,13 | -1.03 | 0.31 | -0.02 | -0.02-0.14 | 0,89 | 9 | 9 | |
| R V Striatum | 17, 5, -2 | 3 -0.01 | -0,07 | 0.08 | 0.08 | 0.08 | 0.50 | 0,08 | 0.66 | 0.51 | 0,11 | 0,11 | 0.92 | 0.36 | 0.36 | |||
| l Муравей INSULA | -24, 10, 7 | -0-02 | -0.22 | 3 0.83 | 0,17 | 1,38 | 0,17 | 0,18 | 1,45 | 0,14 | 0,37 | 3,29 | <0.01 | |||||
| R муравей островковой | 42, 5, 11 | 0.00 | 0,00 | 0,00 | 1.0 | 0,02 | 0,19 | 0,85 | 0,85 | 0,84 | 0,41 | 0,41 | 0,25 2.16 | 0,03 | ||||
| Perig АСС | -1, 37, 16 | -0,04 -0,36 | 0,72 0,08 | 0,65 | 0,52 | -0,04 -0,34 | 0.73 | -0774 | -0.01 | -0.11 | 0,11 | |||||||
| R VOFC | R VOFC | 12, 44, 2 | 0,01 | 0,09 | 0.09 | 0,22 | 3 1.86 | 0,07 | 0,00 | 0,00 | 1,0 | 0,15 | 1,23 | 0,22 | ||||
| 17, 57, 4 | 0,03 | 0,25 | 0,80 | 0,29 | 2.50 | 4 | 0.02 0.02 4 | -0.13 | -1.10 | 0.28 | 0,28 | 0,19 | 1,55 | 0,13 | ||||
| 33, 55, 4 | 0.23 | 1,96 | 0,05 | 0,23 | 1,96 | 0,05 | 0,05 | 0,41 | 0,68 | 0,12 | 0,97 | 0,33 | ||||||
| 28, 53, -2 | -0774 | -0.04-0.32 | 0.75 | 0,75 | 0,17 | 3 1.39 | 3 0,17 | 9074 9073 -0,04-0.31 | 0,76 -0,03 -0,21 | |||||||||
| 35, 46, 18 | 0,03 | 0,24 | 0,81 | -0,06 -0,51 | 0,62 -0,14 | -1.14 | 3 -1.14 | 0,26 | -0.07 | -0.62 | 0.54 | 4 | ||||||
| 28, 46, -4 | -0.09 | — 0,76 | 3 0,45 | 0.24 | 2,04 | 0,05 | 0,15 | 1,23 | 0,22 | 0,22 | 1,89 | 0,06 | ||||||
| л VOFC | -28, 50, 7 | 0,07 | 0,55 | 0.58 | 0.33 | 2 | 3 2,90 4 | <0,01 | -0.01 | -0 0.93 | 0.93 | 3 0,11 | 3 0.87 | 0,39 | | ||||
| -42, 44, | 4 0.12 | 1,00 0,32 | 0,26 | 2,21 | 0,03 | 0,08 | 0,65 | 0,52 | 0.27 2 232 | 0,02 | ||||||||
| -21, 46, -7 | -7 | 9074-0.29 | 0.77 | 0,35 | 3,04 | <0.01 | 0,07 | 0,56 | 0,58 | 0,20 | 1,7 | 0,10 | ||||||
| R ДЛПФК | 33, 14, 45 | -0.13 | -1.05 | -1.050.30 | -0.22 | -1.84 | 0,07 | -0,13 | -1 09 | — 0.30 | -0,19 | 3 -1.61 | 0,11 | |||||
| л ДЛПФК | -37, 17, 36 | -0,01 -0,09 | 0,93 -0,13 -1,07 | 0,29 -0,15 -1,21 | 0,23 | –0,30 | –2,62 | 0,01 | ||||||||||
| 0 | ||||||||
| Пятифакторное решение VOI 1 активность во время нормального состояния кожи | ||||||||
| EigenValue | EigenValue | EigenValue | Кумулятивные (%) | Кумулятивные (%) | регресса: интенсивность | регрессия: неприятность | ||
| 9077 9 | R | P | R | P | ||||
| 1 | 3 3.02 | 30,21 | 30,21 | –0,01 | н.с. 2 | 0,06 | н.с. | |
| 2 | 1,89 | 18,86 | 49,06 | –0,05 | н.с. | 0,06 | н.с. | |
| 3 | 1,37 | 13,67 | 62,73 | –0,01 | н.с. | –0,10 | н.с. | |
| 4 | 1,12 | 11,18 | 73,92 | 0,01 | н.с. | 0,14 | н.с. | |
| 5 | 1,10 | 10,89 | 84,81 | –0,06 | н.с. | –0,18 | н.с. | |
| Трехфакторное решение активности VOI при состоянии кожи, обработанном капсаицином | ||||||||
| 1 | 3.63 | 36,28 36,28 | 0,24 | 0,03 | 0,30 | <0.01 | ||
| 2 1,64 | 16,38 52,66 | -0,33 | <0.01 4 | -0.26 -0.26 4 | <0,01 | |||
| 3 | 1.53 | 15.28 | 3 67.95 | –0,12 | н.с. | -0473 -0.48 4| <0 0.001 | | |
| Пятифакторное решение VOI 1 активность во время нормального состояния кожи | |||||||
| Главный компонент | EigenValue | Диспензия (%) | Накопительное (%) | регрессии: интенсивность | регрессии: неприятности | ||
| г | Р | г | Р | ||||
| 1 | 3.02 | 30,21 | 30,21 | –0,01 | н.с. 2 | 0,06 | н.с. |
| 2 | 1,89 | 18,86 | 49,06 | –0,05 | н.с. | 0,06 | н.с. |
| 3 | 1,37 | 13,67 | 62,73 | –0,01 | н.с. | –0,10 | н.с. |
| 4 | 1,12 | 11,18 | 73,92 | 0,01 | н.с. | 0,14 | н.с. |
| 5 | 1,10 | 10,89 | 84,81 | –0,06 | н.с. | –0,18 | н.с. |
| Трехфакторное решение активности VOI при состоянии кожи, обработанном капсаицином | |||||||
| 1 | 3.63 | 36,28 36,28 | 0,24 | 0,03 | 0,30 | <0.01 | |
| 2 1,64 | 16,38 52,66 | -0,33 | <0.01 4 | -0.26 -0.26 4 | <0,01 | ||
| 3 | 1.53 | 15.28 | 3 67.95 | –0,12 | н.с. | -0473 -0.48 4| <0 0.001 | |
| Пятифакторное решение VOI 1 активность во время нормального состояния кожи | |||||||
| Главный компонент | EigenValue | Диспензия (%) | Накопительное (%) | регрессии: интенсивность | регрессии: неприятности | ||
| г | Р | г | Р | ||||
| 1 | 3.02 | 30,21 | 30,21 | –0,01 | н.с. 2 | 0,06 | н.с. |
| 2 | 1,89 | 18,86 | 49,06 | –0,05 | н.с. | 0,06 | н.с. |
| 3 | 1,37 | 13,67 | 62,73 | –0,01 | н.с. | –0,10 | н.с. |
| 4 | 1,12 | 11,18 | 73,92 | 0,01 | н.с. | 0,14 | н.с. |
| 5 | 1,10 | 10,89 | 84,81 | –0,06 | н.с. | –0,18 | н.с. |
| Трехфакторное решение активности VOI при состоянии кожи, обработанном капсаицином | |||||||
| 1 | 3.63 | 36,28 36,28 | 0,24 | 0,03 | 0,30 | <0.01 | |
| 2 1,64 | 16,38 52,66 | -0,33 | <0.01 4 | -0.26 -0.26 4 | <0,01 | ||
| 3 | 1.53 | 15.28 | 3 67.95 | –0,12 | н.с. | -0473 -0.48 4| <0 0.001 | |
| Пятифакторное решение VOI 1 активность во время нормального состояния кожи | |||||||
| Главный компонент | EigenValue | Диспензия (%) | Накопительное (%) | регрессии: интенсивность | регрессии: неприятности | ||
| г | Р | г | Р | ||||
| 1 | 3.02 | 30,21 | 30,21 | –0,01 | н.с. 2 | 0,06 | н.с. |
| 2 | 1,89 | 18,86 | 49,06 | –0,05 | н.с. | 0,06 | н.с. |
| 3 | 1,37 | 13,67 | 62,73 | –0,01 | н.с. | –0,10 | н.с. |
| 4 | 1,12 | 11,18 | 73,92 | 0,01 | н.с. | 0,14 | н.с. |
| 5 | 1,10 | 10,89 | 84,81 | –0,06 | н.с. | –0,18 | н.с. |
| Трехфакторное решение активности VOI при состоянии кожи, обработанном капсаицином | |||||||
| 1 | 3.63 | 36,28 36,28 | 0,24 | 0,03 | 0,30 | <0.01 | |
| 2 1,64 | 16,38 52,66 | -0,33 | <0.01 4 | -0.26 -0.26 4 | <0,01 | ||
| 3 | 1.53 | 15.28 | 3 67.95 | –0,12 | н.с. | –0,48 | <0,001 |
Ссылки
Andersen E. Околоводопроводная серая кора и кора головного мозга модулируют ответы нейронов медиального таламуса на вредную стимуляцию.
Мозг Res
1986
;375
:30
–6.Бантик С.Дж., Уайз Р.Г., Плогхаус А., Клэр С., Смит С.М., Трейси И. Визуализация того, как внимание модулирует боль у людей, с использованием функциональной МРТ.
Мозг
2002
;125
:310
–9.Барон Р., Барон Ю., Дисброу Э., Робертс Т.П. Мозговая обработка вторичной гипералгезии, вызванной капсаицином: функциональное МРТ-исследование.
Неврология
1999
;53
:548
–57.Басбаум А.И., Филдс Х.Л. Эндогенные механизмы контроля боли: обзор и гипотеза.
Энн Нейрол
1978
;4
:451
–62.Бесерра Л., Брайтер Х.К., Уайз Р., Гонсалес Р.Г., Борсук Д.Поощряйте активацию схем вредными тепловыми раздражителями.
Нейрон
2001
;32
:927
–46.Bornhövd K, Quante M, Glauche V, Bromm B, Weiller C, Büchel C. Болезненные стимулы вызывают различные функции стимул-реакция в миндалевидном теле, префронтальной области, островке и соматосенсорной коре: однократное исследование фМРТ.
Мозг
2002
;125
:1326
–36.Бюхель К., Фристон К.Дж. Модуляция связи в зрительных путях вниманием: корковые взаимодействия, оцененные с помощью моделирования структурных уравнений и фМРТ.
Кора головного мозга
1997
;7
:768
–78.Bunge SA, Ochsner KN, Desmond JE, Glover GH, Gabrieli JDE. Префронтальные области участвуют в хранении информации в памяти и вне ее.
Мозг
2001
;124
:2074
–86.Кейси КЛ. Механизмы переднего мозга ноцицепции и боли: анализ с помощью визуализации.
Proc Natl Acad Sci USA
1999
;96
:7668
–74.Кейси К.Л., Лоренц Дж. Еще раз о детерминантах боли: координаты в сенсорном пространстве.
Управление обезболиванием
2000
;5
:197
–204.Кейси К.Л., Миношима С., Морроу Т.Дж., Кёппе Р.А. Сравнение паттернов активации головного мозга человека при воздействии тепла на кожу, тепловой боли и глубокой холодовой боли.
J Нейрофизиол
1996
;76
:571
–81.Кейси К.Л., Морроу Т.Дж., Лоренц Дж., Миношима С. Временная и пространственная динамика активности переднего мозга человека во время тепловой боли: анализ с помощью позитронно-эмиссионной томографии.
J Нейрофизиол
2001
;85
:951
–9.Каттель РБ. Тест осыпи для количества факторов.
Multivar Behav Res
1966
;1
:245
–76.Когхилл Р.С., Санг К.Н., Майсог Дж.М., Иадарола М.Дж. Обработка интенсивности боли в человеческом мозгу: двусторонний распределенный механизм.
J Нейрофизиол
1999
;82
:1934
–43.Кондес-Лара М., Омана-Запата И., Леон-Олеа М., Санчес-Альварес М.Дорсальный шов и ноцицептивная стимуляция вызывают конвергентные ответы на нейронах центрального латерального таламуса и медиальной префронтальной коры.
Мозг Res
1989
;499
:145
–52.Купер С.Дж. Анестезия префронтальной коры и реакция на вредную стимуляцию.
Природа
1975
;254
:439
–40.Крейг А.Д., Достровский Ю.О. Продолговатый мозг к таламусу. В: Wall PD, Melzack R, редакторы.Учебник боли. 4-е изд. Эдинбург: Черчилль Ливингстон;
1999
. п.183
–214.Крейг А.Д., Чен К., Бэнди Д., Рейман Э.М. Термосенсорная активация островковой коры.
Nat Neurosci
2000
;3
:184
–90.Danziger N, Weil-Fugazza J, Le Bars D, Bouhassira D. Изменение нисходящей модуляции ноцицепции при моноартрите у крыс.
J Neurosci
1999
;19
:2394
–400.Диас Р., Роббинс Т.В., Робертс А.С. Диссоциация в префронтальной коре аффективных сдвигов и переключений внимания.
Природа
1996
;380
:69
–72.Экклстон С., Кромбез Г. Боль требует внимания: когнитивно-аффективная модель прерывающей функции боли. [Обзор].
Psychol Bull
1999
;125
:356
–66.Fox PT, Raichle ME. Зависимость регионарного мозгового кровотока в стриарной коре человека от скорости стимула, продемонстрированная с помощью позитронно-эмиссионной томографии.
J Нейрофизиол
1984
;51
:1109
–20.Фристон К.Дж., Фрит К.Д., Лиддл П.Ф., Фраковяк Р.С. Сравнение функциональных (ПЭТ) изображений: оценка значительных изменений.
J Cereb Blood Flow Metab
1991
;11
:690
–9.Friston KJ, Ungerleider LG, Jezzard P, Turner R. Характеристика модуляторных взаимодействий между V1 и V2 в коре головного мозга человека с помощью фМРТ.
Hum Brain Map
1995
;2
:211
–24.Фунахаши С. Нейрональные механизмы исполнительного контроля префронтальной корой.
Neurosci Res
2001
;39
:147
–65.Gear RW, Алей К.О., Левин Д.Д. Анальгезия, вызванная болью, опосредованная мезолимбическими цепями вознаграждения.
J Neurosci
1999
;19
:7175
–81.Грачев И.Д., Фредриксон Б.Е., Апкарян А.В. Аномальная химия мозга при хронической боли в спине: исследование спектроскопии протонного магнитного резонанса in vivo.
Боль
2000
;89
:7
–18.Грачев И.Д., Томас П.С., Рамачандран Т.С. Снижение уровня N-ацетиласпартата в дорсолатеральной префронтальной коре в случае некупируемой тяжелой симпатически опосредованной хронической боли (комплексный регионарный болевой синдром, тип I).
Brain Cogn
2002
;49
:102
–13.Харди SGP. Проекции на средний мозг медиальной и латеральной префронтальной коры крысы.
Neurosci Lett
1986
;63
:159
–64.Hardy SGP, Haigler HJ. Префронтальные влияния на средний мозг: возможный путь модуляции боли.
Мозг Рес
1985
;339
:285
–93.Хиракава Н., Тершнер С.А., Филдс Х.Л., Мэннинг Б.Х. Двунаправленные изменения аффективного состояния, вызванные манипуляциями с медуллярной схемой модуляции боли.
Неврология
2000
;100
:861
–71.Hsieh J-C, Stahle-Backdahl M, Hagermark O, Stone-Elander S, Rosenquist G, Ingvar M. Травматическая ноцицептивная боль активирует гипоталамус и периакведуктальный серый: исследование позитронно-эмиссионной томографии.
Боль
1996
;64
:303
–14.Ядарола М.Дж., Берман К.Ф., Зеффиро Т.А., Байас-Смит М.Г., Грейсли Р.Х., Макс М.Б. и др. Нервная активация во время острой боли, вызванной капсаицином, и аллодиния, оцененная с помощью ПЭТ.
Мозг
1998
;121
:931
–47.Кунисио К., Хабер С.Н. Цингулостриарная проекция приматов: лимбическая стриарная полоса против сенсомоторной стриарной.
J Comp Neurol
1994
;350
:337
–56.Ле Барс Д., Дикенсон А.Х., Бессон Дж.М. Диффузный вредный ингибирующий контроль (DNIC). I. Воздействие на конвергентные нейроны задних рогов крысы.
Боль
1979
;6
:283
–304.Лоренц Дж., Бромм Б. Потенциальные корреляты вмешательства между когнитивными функциями и тонической экспериментальной болью, связанные с событиями.
Психофизиология
1997
;34
:436
–45.Лоренц Дж., Кросс Д., Миношима С., Морроу Т., Полсон П., Кейси К. Уникальное представление тепловой аллодинии в человеческом мозгу.
Нейрон
2002
;35
:383
–93.Макдональд А.В., Коэн Д.Д., Стенгер В.А., Картер К.С. Диссоциация роли дорсолатеральной префронтальной и передней поясной коры в когнитивном контроле.
Наука
2000
;288
:1835
–8.Мелзак Р. Краткий опросник McGill Pain.
Боль
1987
;30
:191
–7.Мелзак Р., Кейси К.Л. Сенсорные, мотивационные и центральные детерминанты боли. В: Кеншало Д.Р., редактор. Кожа чувствует. Спрингфилд (Иллинойс): Чарльз С. Томас;
1968
. п.423
–43.Мелзак Р., Уолл, полиция. Болевые механизмы: новая теория.
Наука
1965
;150
:971
–9.Миношима С., Кёппе Р.А., Фрей К.А., Куль Д.Е. Анатомическая стандартизация: линейное масштабирование и нелинейное искажение функциональных изображений мозга.
J Nucl Med
1994
;35
:1528
–37.Наута WJH, Кайперс HGJM. Некоторые восходящие пути в ретикулярной формации ствола головного мозга кошки. В Jasper HH, Proctor LD, редакторы. Ретикулярная формация головного мозга. Бостон: Литтл, Браун;
1958
. п.3
–30.Northoff G, Richter A, Gessner M, Schlagenhauf F, Fell J, Baumgart F, et al.Функциональная диссоциация между медиальной и латеральной пространственно-временной активацией префронтальной коры при отрицательных и положительных эмоциях: комбинированное исследование фМРТ/МЭГ.
Кора головного мозга
2000
;10
:93
–107.Пасториза Л.Н., Морроу Т.Дж., Кейси К.Л. Поражения медиальной лобной коры избирательно ослабляют реакцию горячей пластинки: возможная ноцифенсивная апраксия у крыс.
Боль
1996
;64
:11
–7.Полсон Ч.П., Миношима С., Морроу Т.Дж., Кейси К.Л.Гендерные различия в восприятии боли и паттернах активации головного мозга при вредной тепловой стимуляции у людей.
Боль
1998
;76
:223
–9.Петрович П., Петерссон К.М., Гатан П.Х., Стоун-Эландер С., Ингвар М. Активация головного мозга, связанная с болью, изменяется отвлекающей когнитивной задачей.
Боль
2000
;85
:19
–30.Петрович П., Калсо Э., Петерссон К.М., Ингвар М. Плацебо и опиоидная анальгезия — визуализация общей нейронной сети.
Наука
2002
;295
:1737
–40.Peyron R, Garcia-Larrea L, Groegoire MC, Costes N, Convers P, Lavenne F, et al. Гемодинамические реакции мозга на острую боль у людей. Сенсорные сети и сети внимания.
Мозг
1999
;122
:1765
–80.Плогхаус А., Трейси И., Гати Дж.С., Клэр С., Менон Р.С., Мэтьюз П.М. и др. Диссоциация боли от ее ожидания в человеческом мозгу.
Наука
1999
;284
:1979
–81.Плогхаус А., Нараин С., Бекманн С.Ф., Клэр С., Бантик С., Уайз Р. и др. Обострение боли при тревоге связано с активностью в сети гиппокампа.
J Neurosci
2001
;21
:9896
–903.Цена ДД. Психологические и нервные механизмы аффективного измерения боли.
Наука
2000
;288
:1769
–72.Raichle ME, MacLeod AM, Snyder AZ, Powers WJ, Gusnard DA, Shulman GL.Режим работы мозга по умолчанию.
Proc Natl Acad Sci USA
2001
;98
:676
–82.Рейнвилл П., Дункан Г.Х., Прайс Д.Д., Перевозчик Б., Бушнелл М.С. Болевой аффект закодирован в передней части поясной извилины человека, но не в соматосенсорной коре.
Наука
1997
;277
:968
–71.Решетняк В.К., Кукушкин М.Л. Влияние удаления орбитофронтальной коры на развитие рефлекторной анальгезии. [Русский].
Биул Эксп Биол Мед
1989
;108
:14
–6.Rosen SD, Paulesu E, Nihoyannopoulos P, Tousoulis D, Frackowiak RSJ, Frith CD, et al. Тихая ишемия как центральная проблема: региональная активация мозга по сравнению с тихой и болевой ишемией миокарда.
Энн Интерн Мед
1996
;124
:939
–49.Сакаи К., Роу Дж. Б., Пассингем Р.Э. Активное обслуживание в префронтальной области 46 создает память, устойчивую к дистракторам.
Nat Neurosci
2002
;5
:479
–84.Салинас Э., Сейновски Т.Дж. Коррелированная активность нейронов и поток нейронной информации.
Nat Rev Neurosci
2001
;2
:539
–50.Савамото Н., Хонда М., Окада Т., Ханакава Т., Канда М., Фукуяма Х. и др. Ожидание боли усиливает реакцию на безболевую соматосенсорную стимуляцию в передней части поясной извилины и теменной покрышке/задней доле: исследование функциональной магнитно-резонансной томографии, связанное с событием.
J Neurosci
2000
;20
:7438
–45.Сильверман Д.Х., Мунаката Дж., Эммес Х., Манделькерн М.А., Хох К.К., Майер Э.А. Регионарная мозговая активность при нормальном и патологическом восприятии висцеральной боли.
Гастроэнтерология
1997
;112
:64
–72.Smith EE, Jonides J. Хранение и исполнительные процессы в лобных долях.
Наука
1999
;283
:1657
–61.Tölle TR, KaufmannT, Siessmeier T, Lautenbacher S, Berthele A, Munz F, et al. Регионально-специфическое кодирование сенсорных и аффективных компонентов боли в мозге человека: корреляционный анализ позитронно-эмиссионной томографии.
Энн Нейрол
1999
;45
:40
–7.Уорсли К.Дж., Эванс А.С., Марретт С., Нилин П. Трехмерный статистический анализ исследований активации CBF в мозге человека.
J Cereb Blood Flow Metab
1992
;12
:900
–18.Чжан С., Тан Дж.-С., Юань Б., Цзя Х. Ингибирующие эффекты электрической стимуляции вентролатеральной орбитальной коры на рефлекс открывания челюсти у крыс.
Мозг Рес
1998
;813
:359
–66.Различные вклады вискерной сенсорной коры и языко-челюстной моторной коры в целенаправленную сенсомоторную трансформацию
https://doi.org/10.1016/j.neuron.2019.07.008Get rights and content Локализация язычно-челюстной моторной коры с помощью анатомо-функционального картированияИнактивация язычно-челюстной моторной коры тормозит целенаправленное контралатеральное облизывание
не сенсорный сигнал
Нейроны в сенсорной коре усов кодируют сенсорный сигнал, но не направление облизывания
Резюме
Нейронные цепи, лежащие в основе целенаправленных сенсомоторных преобразований в мозге млекопитающих, изучены не полностью.Здесь мы сравнили роль первичной язычно-челюстной моторной коры (tjM1) и первичной сенсорной коры усов (wS1) у мышей с ограниченной головой, обученных облизывать носик вознаграждения в ответ на отклонение усов. Двухфотонная микроскопия в сочетании с микропризмами позволила визуализировать активность нейронной сети в слоях коры у трансгенных мышей, экспрессирующих генетически кодируемый индикатор кальция. Ранняя фаза активности в wS1 кодировала сенсорный стимул усов и была необходима для обнаружения стимулов усов.Активность в tjM1 кодировала направление лизания во время выполнения задания и была необходима для контралатерального лизания. Предстимульная активность в tjM1, но не в wS1, предсказывала направление лизания и причинно способствовала небольшим подготовительным движениям челюстей. Наши данные показывают сдвиг в схеме кодирования от wS1 к tjM1, что согласуется с гипотезой о том, что эти области представляют собой корковые точки начала и окончания для этой целенаправленной сенсомоторной трансформации.
Ключевые слова
Ключевые слова
Ключевые слова
Назначение назовавших назовательство
Цель-направленное сенсорное преобразование
усы Сенсорная обработка
Соматосенсорные Cortex
Облизывание
Мотор Cortex
Рекомендуемые статьи Статьи (0)
© 2019 Автор (ы).Опубликовано Elsevier Inc.
Рекомендуемые статьи
Ссылки на статьи
CRS0010110883.TIF
%PDF-1.4 % 159 0 объект > эндообъект 155 0 объект [/КалГрей>] эндообъект 154 0 объект [/КалРГБ>] эндообъект 156 0 объект >поток Пт Сен 07 17:19:33 2007PageGenie PDFGeneratorapplication/pdf
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка браузера на прием файлов cookie
Существует множество причин, по которым файл cookie не может быть установлен правильно. Ниже приведены наиболее распространенные причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки браузера, чтобы принять файлы cookie, или спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файл cookie.
- Ваш браузер не поддерживает файлы cookie. Попробуйте другой браузер, если вы подозреваете это.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы это исправить, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Предоставить доступ без файлов cookie потребует от сайта создания нового сеанса для каждой посещаемой вами страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в файле cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, если вы не решите ввести его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступ к остальной части вашего компьютера, и только сайт, создавший файл cookie, может его прочитать.
Кортексин-2 человека (CTXN2) Набор ИФА
Цена: 797,50 долларов США(Размер: 96 тестов)
Нажмите на изображение, чтобы увидеть легенду изображения
Набор ИФА человеческого кортексина-2 (CTXN2) представляет собой набор ИФА для количественного измерения in vitro концентраций человеческого кортексина-2 (CTXN2) в гомогенатах тканей, клеточных лизатах и других биологических жидкостях.| Цель | Кортексин-2 (CTXN2) |
| Реактивность | Человек |
| Протестированные приложения | ИФА |
| Рекомендуемые разведения | Оптимальные разведения/концентрации должны определяться конечным пользователем. |
| Хранение | Поставляется при температуре 4 °C. После получения храните набор в соответствии с инструкциями по хранению в руководстве к набору. |
| Срок действия | Срок действия данного комплекта 6 месяцев. |
| Устойчивость | Стабильность набора определяется скоростью потери активности. Потери составляют менее 5% в течение срока годности при соответствующих условиях хранения. Чтобы свести к минимуму колебания производительности, необходимо строго контролировать рабочие процедуры и лабораторные условия. Также настоятельно рекомендуется, чтобы весь анализ выполнялся одним и тем же пользователем. |
| Первичный преобразователь частоты UniProt | P0C2S0 ( ЮниПрот , ExPASy ) |
| Имя записи UniProt | CTXN2_ЧЕЛОВЕК |
| Символ гена | CTXN2 |
| GeneID | 399697 |
| ХГНК | 31109 |
| БОЧКА | ВСА: 399697 |
| Испытательный полигон | 0.156 нг/мл — 10 нг/мл |
| Стандартная форма | Лиофилизированный |
| Метод обнаружения | Колориметрический |
| Данные анализа | Количественный |
| Тип образца | Гомогенаты тканей, лизаты клеток и другие биологические жидкости. |
| Наличие | Отправка в течение 5-15 рабочих дней. |
| Примечание | Этот продукт предназначен только для исследовательских целей. Диапазон и чувствительность могут быть изменены. Пожалуйста, свяжитесь с нами для получения последней информации о продукте.Для получения точных результатов концентрации пробы должны быть разбавлены до средней концентрации набора. Если вам требуется определенный ассортимент, пожалуйста, свяжитесь с нами заранее или напишите свой запрос в комментариях к заказу. Обратите внимание, что наши наборы ELISA и CLIA оптимизированы для обнаружения нативных образцов, а не рекомбинантных белков. Мы не можем гарантировать обнаружение рекомбинантных белков, поскольку они могут иметь другую последовательность или третичную структуру по сравнению с нативным белком. |
Различная функция рецепторов эстрогена в передней поясной коре грызунов при отвращении к боли | Анестезиология
Крысы Sprague-Dawley (взрослые: от 2 до 3 месяцев, от 200 до 220 г; молодые: от 2 до 3 недель, от 50 до 80 г, только для электрофизиологических исследований) и мыши C57BL/6 (от 8 до 12 недель, 20 до 25 g) были получены из Шанхайского центра экспериментальных животных Китайской академии наук, Шанхай, Китай.Мышей CaMK2α::EYFP (в возрасте от 8 до 12 недель, от 20 до 25 г) разводили путем скрещивания мышей CaMK2α-Cre (каталожный № 005359 от Jackson Laboratory, США) и мышей R26R-EYFP (каталожный № 006148 от Jackson Laboratory). . В общей сложности 300 самцов и самок крыс и 46 самцов мышей использовали для исследований in vivo и исследований in vitro (таблица 1). Всех животных содержали при 12:12-часовом цикле свет-темнота в помещении с кондиционированием воздуха (23 ± 2°C) с водой и пищей ad libitum . Животных нумеровали, случайным образом распределяли по разным экспериментальным группам, а затем тестировали в последовательном порядке.Все эксперименты на животных были одобрены Комитетом по использованию экспериментов на животных Университета Фудань (разрешение № SYXK 2009-0082) и соответствовали политике использования лабораторных животных, изданной Международной ассоциацией по изучению боли (Вашингтон, округ Колумбия). ). После экспериментов животных подвергли эвтаназии путем ингаляции углекислого газа. Все поведенческие тесты, электрофизиологические записи, а также Вестерн-блоттинг и иммуногистохимические эксперименты, описанные здесь, были выполнены экспериментаторами, которые не знали о методах лечения.Первичным результатом было влияние фармакологических и генетических вмешательств с участием различных рецепторов эстрогена в ростральной передней части поясной извилины на отвращение, связанное с болью. Анальгетики в этом исследовании не использовались.
1,3-Бис(4-гидроксифенил)-4-метил-5-[4-(2-пиперидинилэтокси)фенол]-1H-пиразолдигидрохлорид (MPP) и PHTPP были приобретены у Tocris Bioscience (Великобритания). (3aS*,4R*,9bR*)-4-(6-бром-1,3-бензодиоксол-5-ил)-3a,4,5,9b-3H-циклопента[c]хинолон (G15), 4, 4´,4´´-(4-пропил-[1H]-пиразол-1,3,5-триил)трисфенол (ППТ), 2,3-бис(4-гидроксифенил)-пропионитрил (ДПН), (±) -1-[(3aR*,4S*,9bS*)-4-(6-бром-1,3-бензодиоксол-5-ил)-3a,4,5,9b-тетрагидро-3H-циклопента[c]хинолин -8-ил]-этанон, 17β-эстрадиол (основная форма эстрогена), Rp–аденозин-3´,5´-циклический монофосфотиотиотиотиотиотиотиотиотриэтиаммониевой соли гидрат (цАМФ), (1S,6bR,9aS, 11R,11bR) 11 -(ацетилокси)-1,6b,7,8,9а,10,11,11b-октагидро-1-(метоксиметил)-9а,11b-диметил-3Н-фуро(4,3,2-де)индено(4 ,5,-h)-2-)]-2-бензопиран-3,6,9-трион (вортманнин), кунжутное масло и диметилсульфоксид были приобретены у Sigma (США).